6. Восприятие яркости
Известно, что существует примитивное скотоводческое племя, в языке которого нет слова «зеленый», однако есть шесть слов для обозначения разных оттенков красного. Люди этого племени, занимающиеся различными ремеслами, вкладывают в каждое из них специфическое значение. Прежде чем перейти к рассмотрению проблемы восприятия яркости и цвета, мы остановимся на минуту, чтобы уточнить некоторые понятия, подобно тому как плотник не приступит к работе, пока не отточит свои инструменты.
Мы говорим об интенсивности света, раздражающего глаз; это качество светового раздражителя дает ощущение яркости. Интенсивность — это физическая энергия света, которая может быть измерена с помощью различного рода фотометров, включая хорошо известный фотографам экспонометр. Яркость — это субъективное ощущение интенсивности света. Мы уверены, что знаем, что имеет в в виду человек, говоря: «Какой яркий день!» Он имеет в виду не только то, что он может фотографировать на малочувствительную пленку, но также и то, что он испытывает слепящее ощущение. Это ощущение — только грубое отражение интенсивности света, доходящего до глаза.
Когда мы говорим о восприятии цвета, мы, собственно, говорим не столько о цветах, сколько об оттенках. Это делается попросту для того, чтобы избежать трудности, так как в самом деле под «цветами» мы имеем в виду те ощущения, которые могут быть обозначены словом «красный» или «синий». Таким образом, специалисты говорят скорее о «спектральных оттенках», чем о «спектральных цветах», однако в этом не всегда есть необходимость. Гораздо важнее различие между интенсивностью ж яркостью.
Другое важное различие, которое следует делать, это различие между цветом как ощущением и цветом как длиной волны (или группой длин волн), доходящей до глаза. Строго говоря, свет сам по себе не окрашен: он вызывает ощущение яркости и цвета, однако только при наличии соответствующего глаза и нервной системы. Язык специалистов смешивает эти понятия: мы говорим иногда об «окрашенном свете», например, о «желтом свете», хотя это неточно. Правильнее было бы говорить о свете, который обычно вызывает ощущение, обозначаемое большинством людей как «желтое».
Не пытаясь объяснить, каким образом физические качества — интенсивность и длина волны света — вызывают различные ощущения (в конечном счете мы еще не знаем, как ответить на этот вопрос), мы должны отчетливо представить себе, что без соответствующей нервной системы не было бы яркости и цвета. Пока не возникла жизнь, все было безмолвно, хотя горы рушились.
Наиболее простым из зрительных ощущений является ощущение яркости. Нельзя описать это ощущение. Слепой человек ничего не знает о нем, но для остальных людей мир создается через яркость и цвет. Противоположное ощущение темноты такое же сильное — мы говорим о «плотной стене темноты, которая давит на нас», однако для слепого это ощущение просто не существует. Ощущение, возникающее при отсутствии света, и есть ощущение темноты, слепой же полностью лишен зрительных ощущений. Мы ближе всего подошли бы к описанию мира слепых, у которых нет ощущения яркости и темноты, представив себе то, что находится позади нашей головы. Мы не ощущаем темноты позади нас, мы не ощущаем ничего, а это совсем другое.
Яркость не просто ощущение интенсивности света, раздражающего сетчатку. Ощущение яркости, возникающее при данной интенсивности, зависит от степени адаптации глаза, а также от целого ряда различных сложных условий, определяющих явление контраста объектов пли пятен света. Иными словами, яркость — это функция не только интенсивности света, попадающего на определенный участок сетчатки в данный момент, но также функция интенсивности света, который возбуждал сетчатку в недавнее время, как и функция интенсивности света, падающего на другие участки сетчатки.

Рис. 6, 1. Явление одновременного контраста. Та часть серого круга, которая находится на черном фоне, кажется несколько светлее, чем другая, расположенная на белом фоне. Этот эффект усиливается, если на границе белого и черного фона на круг положить тонкую нить.
АДАПТАЦИЯ К СВЕТУ И ТЕМНОТЕ
Если глаз находится некоторое время в темноте, он становится более чувствительным, и данное освещение начинает казаться более ярким. Эта так называемая темновая адаптация возникает в течение первых нескольких минут пребывания в темноте. Палочковые и колбочковые рецепторные клетки адаптируются с различной быстротой: адаптация колбочек завершается в пределах семи минут, в то время как адаптация палочек продолжается в течение часа или больше. Это можно видеть на рис. 6, 2, который показывает, что существуют две адаптационные кривые: одна — для палочек, другая — для колбочек. Можно сказать, что в глазу имеются две переплетающиеся друг с другом сетчатки.
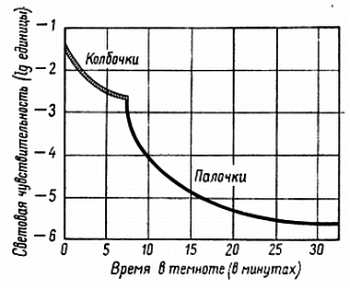
Рис. 6, 2. Увеличение чувствительности глаза в темноте, известное под названием темновая адаптация. Заштрихованная кривая показывает ход адаптации колбочек, черная — ход адаптации палочек; последняя протекает медленнее, но приводит к большей чувствительности глаза. При тусклом освещении функционируют только палочки, в то время как при ярком свете, когда активны колбочки, они, вероятно, затормаживаются.
Механизмы темновой адаптации становятся более понятными благодаря остроумным и технически блестящим экспериментам английского физиолога Раштона. В течение многих лет предполагалось, что адаптация является результатом регенерации зрительного пигмента глаза, который «обесцвечивается» при воздействии света; это «обесцвечивание» каким-то неизвестным пока еще образом вызывает стимуляцию фоторецепторов, после чего электрический сигнал передается в зрительный нерв. Фотохимический родопсин был извлечен из глаза лягушки, и его плотность при воздействии света была измерена во время «обесцвечивания» и при регенерации. Эти данные были сопоставлены с кривыми темновой адаптации человеческого глаза, которые приведены на рис. 6, 2. И действительно, они почти совпадают друг с другом, что указывает на существование тесной связи между фотохимией родопсина и изменяющейся чувствительностью палочкового аппарата глаз. По-видимому, яркость ощущения должна быть связана с количеством фотохимического родопсина, «обесцвечиваемого» под воздействием света. Смысл работы Раштона состоит в том, что он произвел измерение плотности фотохимического родопсина непосредственно в живом глазе во время темновой адаптации или во время воздействия какого-либо окрашенного света, который он применял в опыте. В сущности, методика этого опыта заключается в том, что глазу предъявляется короткая вспышка света и с помощью высокочувствительного фотоэлемента измеряется количество света, отраженного от глаза. Сначала казалось невозможным сделать это с человеческим глазом, так мала масса отраженного света в связи с почти полной абсорбцией света фотохимическими элементами ы черным пигментом, расположенным позади рецептором; поэтому экспериментатор использовал глаз кошки; задний отражающий слой сетчатки (tapetum) служил зеркалом для отражения света на фотоэлемент. Этот метод оправдал себя в эксперименте на кошачьем глазе, и Раштону затем удалось так его усовершенствовать, что он стал достаточно чувствительным, чтобы улавливать и измерять очень слабый свет, отражаемый от человеческого глаза. Он нашел, что по мере адаптации происходит «обесцвечивание» фотохимического вещества, причем отношение между энергией света и массой обесцвечиваемого фотохимического вещества выражается логарифмической зависимостью. Таким образом, он открыл механизм действия светочувствительного пигмента.
КОНТРАСТ
Другим фактором, определяющим ощущение яркости, является интенсивность освещения окружающего поля. Данная поверхность обычно кажется более яркой, если ее окружение темное, а данный цвет воспринимается как более интенсивный, если его окружает поле, окрашенное в дополнительный цвет. Этот факт, бесспорно, связан с процессами взаимодействия рецепторов. Усиление контраста, по-видимому, связано с общим перцептивным фактором — влиянием границ на восприятие объекта. Вероятно, в первую очередь в мозг поступает именно информация о пограничных участках, в то время как области с постоянной освещенностью малоинформативны. Зрительная система экстраполирует информацию об объекте, заключенном в определенные границы, что бесспорно экономит большое количество информации, которое поступает от периферической части системы, хотя и за счет некоторого усложнения процессов, происходящих в высших отделах мозга. Этот процесс известен как латеральное торможение. Несмотря на то что явление контраста и усиления восприятия окружающих объект участков является главным образом результатом действия сетчаточных механизмов, оно определяется, очевидно, также и центральными процессами. Это видно из рис. 6, 1, который демонстрирует отчетливое явление контраста. Окрашенное в один и тот же серый цвет, кольцо кажется более светлым на темном фоне, чем на белом. Этот эффект значительно усиливается, если круг и фон разделяет четкая граница; контраст сильнее, если фигура интерпретируется как состоящая из двух половин, чем если она воспринимается как одно целое. Эти факты указывают на участие центральных мозговых факторов в этом явлении.
Некоторое представление о сложности организации системы восприятия яркости у человека дает парадокс Фехнера. Он состоит в следующем. Если глазу предъявляется маленький, довольно яркий источник света, он будет вызывать ощущение определенной яркости, и зрачок при включении этого источника света будет уменьшаться до определенного размера. Теперь добавим второй, более тусклый источник света. Он помещается несколько в стороне от первого, так, чтобы возбуждать другую область сетчатки. Что же при этом произойдет? Хотя общая интенсивность света с добавлением второго источника увеличится, зрачок больше не сократится, как это можно было бы ожидать, а расширится соответственно разнице интенсивностей между первым и вторым раздражителями. По-видимому, он реагирует не на общее, а на среднее освещение. Никто не знает, как сетчатка осуществляет это.
Попробуем закрыть один глаз и проследить изменения в яркости. Практически нет разницы, воспринимается ли свет одним или двумя глазами. Однако это не так; когда маленькие тусклые источники света воспринимаются в окружающей темноте, тогда они кажутся значительно ярче при работе двух глаз, чем при работе одного. Это явление еще не разгадано.
Рис. 6, 4. Химические процессы, лежащие в основе зрения. Черная кривая показывает чувствительность человеческого глаза (темно-адаптированного) к различной длине световой волны. Красные точки показывают количество света в пределах того же самого диапазона световых волн, которое поглощается фотохимическим родопсином в глазу лягушки. Обе кривые, по существу, совпадают, и это говорит о том, что человеческий глаз при темновой адаптации функционирует путем поглощения света тем же фотохимическим веществом.
Яркость — функция цвета. Когда глаз воспринимает лучи света различного цвета, но одной и той же интенсивности, то цвета, расположенные в середине спектра, будут казаться ярче, чем цвета, расположенные на концах спектра. Это показано на рис. 6, 5; кривая, изображенная на этом рисунке, известна как кривая спектральной яркости света. Это явление имеет практическое значение, так как, если мы хотим, чтобы сигнализирующий об опасности свет был ясно виден, он должен быть окрашен в цвет, к которому глаз максимально чувствителен, то есть в цвет, расположенный в середине спектра. Дело осложняется еще и тем, что кривые чувствительности для палочек и колбочек несколько различны. Они сходны по общему виду, однако колбочки более чувствительны к оранжевому цвету, а палочки — к зеленому. (На этом основании есть смысл окрашивать стены затемненной фотографической комнаты в зеленый цвет, так как глаза при этом получают наиболее эффективный свет, к которому фотографическая пленка относительно нечувствительна.)
Рис. 6, 5. Этот рисунок показывает, как изменяется чувствительность глаза к различной длине световых волн в спектре, когда глаз адаптирован к свету. Черная кривая показывает чувствительность темно-адаптированного глаза, красная кривая показывает, что при адаптации к свету происходит изменение чувствительности к цветам спектра, в это время колбочки берут верх над палочками. Это явление известно под названием «сдвиг Пуркинье».
Кривая яркости света ничего не говорит нам больше о восприятии цвета. Она отражает чувствительность к свету в зависимости от длины световой волны, но вне связи с теми цветами, которые видит глаз при каждой длине световой волны. Глаза животных, не имеющих цветного зрения, обнаруживают сходную с человеческой кривую спектральной яркости света.
Можно предположить, что, помимо фотохимических изменений, связанных с процессом адаптации к свету при восприятии света действуют еще некоторые дополнительные механизмы, причем не фотохимической, а нервной природы. В частности, после завершения процесса адаптации глаза к темноте пространственные и временные характеристики остроты зрения ухудшаются, в то время как чувствительность возрастает. Однако при темновой адаптации утрачивается способность глаза различать мелкие детали. Это непростое явление, оно возникает отчасти вследствие того, что сетчатка интегрирует при этом энергию с большей зоны, то есть от большего числа рецепторных элементов. По ходу темновой адаптации увеличивается время, в течение которого может интегрироваться световая энергия, попадающая на сетчатку.
Изменения временных характеристик чувствительности глаза при темновой адаптации лучше всего, хотя и не в прямой форме, проявляются в любопытном и очень интересном явлении, известном под названием эффект маятника Пульфриха. Не менее примечательна история открытия этого эффекта, особенность которого состоит в том, что его можно наблюдать, только смотря обоими глазами, — и все же он был открыт человеком, слепым на один глаз! Этот эксперимент заслуживает того, чтобы его повторить. Возьмите длинную нитку, прикрепите к ней гирю, чтобы сделать маятник длиной в несколько футов (один фут равен 30, 48 см). Качните маятник под прямым углом к линии взора. Смотрите на колеблющуюся гирю обоими глазами, но прикройте один глаз темным, проницаемым для света стеклом (например, половинкой солнечных очков или кусочком засвеченной пленки). Тогда можно будет видеть, что гиря качается не по прямой линии, а описывает эллипс. Этот эллипс может быть очень странным: в самом деле, длинная ось может располагаться вдоль линии взора и, несмотря на это, будет казаться, что гиря, качаясь по прямой, пересекает эту линию.
Что же вызывает этот удивительный эффект? Уменьшая приток света, темное стекло вызывает процесс темновой адаптации в глазу. Адаптация приводит к задержке передачи сигнала от этого глаза к мозгу; другой глаз не участвует в этом процессе. Эта отсрочка ведет к тому, что затемненный глаз видит гирю с некоторым запозданием, а так как движение гири в середине траектории ускоряется, отсрочка в этом месте оказывается более значительной, и глаз, прикрытый фильтром, видит гирю все дальше и дальше от того места, где видит гирю другой незатемненный глаз. Эта разница в восприятии положения гири одним и другим глазом и приводит к тому, что траектория движения гири кажется эллипсом, расположенным по прямой линии к линии взора; мозг оценивает движение гири как действительно происходящее по эллипсу. Это показано на рис. 6, 3.
Рис. 6, 3. Маятник Пульфриха. Маятник колеблется под прямым углом к линии взора наблюдателя, один глаз которого закрыт темным стеклом, причем оба глаза открыты. Наблюдателю кажется, что маятник описывает эллипс. Этот эффект возникает в результате задержки сигнала от частично адаптированного к темноте глаза, закрытого темным стеклом. При приближении маятника к середине траектории колебания увеличивается разобщение изображений, получаемых левым и правым глазами, которое оценивается мозгом как различие в расстоянии. Это и создает видимость эллипса.
По-видимому, увеличение отсрочки при темновой адаптации связано с увеличением времени интеграции возбуждения, подобно тому как фотограф прибегает к более длительной экспозиции при тусклом освещении. Мы видим этот эффект непосредственно, когда наблюдаем, как увеличивается огненный след, оставляемый в темном небе разорвавшимися ракетами фейерверка, так как в темноте усиливаются процессы темповой адаптации.
Как увеличение задержки передачи сигнала от сетчатки к мозгу, так и связанное с этим процессом увеличение времени интеграции возбуждения, имеет практическое значение. Задержка сигналов сетчатки вызывает увеличение времени реакции у шоферов при тусклом освещении, а увеличение времени интеграции возбуждения ухудшает точную локализацию движущихся объектов. В этих условиях затрудняются игры, связанные с движением; судья провозглашает: «Прекратите игру из-за слабого освещения» — задолго до того, как зрители сами убеждаются в том, что солнце садится.
ЧУВСТВИТЕЛЬНОСТЬ ГЛАЗА К СВЕТУ
При увеличении интенсивности света учащаются импульсы, идущие от рецепторов сетчатки, причем интенсивность света выражается в частоте импульсов. К сожалению, невозможно зарегистрировать электрическую активность рецепторов глаза позвоночных, потому что у них сетчатка «вывернута наизнанку», так что электроды не могут достичь рецепторов без больших повреждений. К тому времени, когда импульсы достигают зрительного нерва, они усложняются благодаря взаимосвязям нервных клеток, расположенных в различных слоях сетчатки.
Существует, однако, такой глаз, в котором рецепторы непосредственно связаны с отдельными нервными волокнами: это глаз своего рода живого ископаемого, краба Limulus, который живет на восточном побережье США. Отдельные нервные волокна глаза этого древнего краба оказались наиболее пригодными для проведения исследования, что, однако, трудно было предположить заранее. Рис. 6, 6 показывает электрическую активность нервного волокна глаза Limulus’a.
Рис. 6, 6. Электрическая активность отдельного волокна зрительного нерва Limulus’a в ответ на три интенсивности света, записанная на осциллоскопе. Частота импульсов увеличивается в соответствии с логарифмом интенсивности света.
Рис. 6, 7. Частота импульсов после различных по длительности периодов темноты. При усилении темновой адаптации частота импульсов возрастает, соответственно увеличивается и ощущение яркости, хотя реальная интенсивность света одна и та же.
Было обнаружено, что в рецепторных клетках глаза этого краба частота импульсов связана приблизительно логарифмической зависимостью с интенсивностью света. Это видно из рис. 6, 8.
Рис. 6, 8. График, построенный на основании записей, приведенных на рисунках 6,6 и 6,7. Частота импульсов сопоставлена с логарифмом интенсивности света. Полученная зависимость выражается почти прямой линией, что говорит о существовании логарифмической зависимости между частотой импульсов и интенсивностью света (при неизменной адаптации).
Первая кривая (рис. 6, 6) показывает низкую частоту импульсов после одноминутной темновой адаптации глаза. На другой кривой (рис. 6, 7) видно, что частота импульсов увеличивается, когда глаз находится в темноте более длительное время. Это соответствует нашему собственному ощущению увеличения яркости света после темновой адаптации.
Что происходит, когда мы смотрим на источник очень слабого света в темной комнате? Можно было бы думать, что, если нет света, отсутствует и активность, передающаяся от сетчатки в мозг; когда же появляется какой-нибудь свет, сетчатка сигнализирует о нем и мы видим свет. Однако дело обстоит не так просто. При полном отсутствии света сетчатка и зрительный нерв не являются полностью инактивными. В них всегда имеется некоторая остаточная нервная активность, которая доходит до мозга, даже если отсутствует какая-либо стимуляция глаза светом. Об этом говорит непосредственная регистрация активности зрительного нерва глаза кошки, полностью адаптированного к темноте, и мы имеем все основания предположить, что это справедливо и по отношению к глазу человека и других животных.
Этот факт постоянного фона спонтанной активности имеет большое значение. Глаз удивительно чувствителен, мы можем видеть вспышку света столь незначительную, что ее трудно зарегистрировать каким-либо искусственным прибором. Однако глаз был бы еще более чувствителен, если бы не было спонтанной активности зрительной системы, которая представляет собой постоянную проблему для мозга.
Представим себе нервные импульсы, приходящие в мозг; являются ли они результатом воздействия света на глаз или они являются просто спонтанным «шумом» зрительной системы? Проблема, стоящая перед мозгом, заключается в том, чтобы «решить», отражает ли эта нервная активность внешнее раздражение или это только «шум», который следует игнорировать. Эта ситуация очень хорошо знакома инженерам связи, потому что во всех чувствительных детекторах может возникать шум — случайная генерация сигналов, которые всегда ухудшают чувствительность детекторов. Существуют способы уменьшения вредного влияния шума; они с успехом применяются в радиоастрономии и при обнаружении слабых земмлетрясений; шум маскирует источники радиоволн в космосе и на Земле, подобно тому как маскируются слабые зрительные сигналы. Глаз использует некоторые приспособления, уменьшающие влияние «шума» и значительно повышающие длительность периода, в течение которого происходит интеграция сигнала, — действие этого механизма мы видели в эффекте Пульфриха, — путем запроса дополнительных подтверждающих сигналов от отдельных рецепторов, которые выступают в качестве независимых свидетелей.
Одним из самых старых законов экспериментальной психологии является закон Вебера. Согласно ему, наименьшее различие интенсивностей, которое может быть воспринято, прямо пропорционально исходной интенсивности света. Например, если в ярко освещенную комнату вносится одна горящая свеча, увеличение освещения едва различимо, но если комната освещена плохо, — скажем, в ней горит только несколько свечей, — тогда добавление еще одной свечи дает заметное увеличение освещенности. Фактически мы можем различать изменение интенсивности, равное примерно одному проценту по отношению к исходной освещенности. Это выражается в формуле ΔI/ΔI = const (где Δ означает минимальную добавку интенсивности к исходной интенсивности I). Этот закон полностью справедлив в отношении довольно широкого диапазона исходных интенсивностей, но он неприменим в случаях малой исходной интенсивности. Это можно видеть из рис. 6, 9, где — если бы закон Вебера был справедлив и при нулевой интенсивности — мы имели бы прямую горизонтальную линию, указывающую на неизменность (инвариантность) едва заметных различий в интенсивности ΔI/I по отношению ко всем исходным значениям I. Фактически мы получаем кривую, которая изображена на этом графике, указывающую, что величина ΔI/I значительно увеличивается, когда исходная интенсивность I света становится небольшой. Это нарушение закона Вебера объясняется главным образом тем, что имеются некоторые остаточные разряды возбуждения клеток сетчатки даже при отсутствии света. Эта остаточная активность для мозга эквивалентна более или менее постоянной слабой освещенности, которая добавляется к исходной. Мы можем оценить ее величину, экстраполируя кривую за пределы оси Y и считывая значения этого графика. Это выражает уровень шума в единицах интенсивности света.
Рис. 6, 9. Закон Вебера (ΔI/I = const). Отношение ΔI к I выражается в виде горизонтальной прямой линии для широкого диапазона значений I, но эта зависимость нарушается в зоне слабых интенсивностей, когда значения ΔI/I должны быть увеличены, чтобы сигнал мог быть выделен из шума. Отношение ΔI к I, по существу, выражается в виде прямой линии вплоть до небольших значений I, что указывает на наличие скрытой константы к в знаменателе. Таким образом, мы можем записать закон Вебера следующим образом: ΔI/I + k = const, где k, по-видимому, связана с уровнем шума в нервной системе. Этот уровень увеличивается с возрастом.
Скрытая константа k может быть отнесена за счет «шума» сетчатки. Существуют доказательства того, что этот внутренний шум зрительной системы увеличивается с возрастом: повышение уровня шума, безусловно, является отчасти причиной постепенного ухудшения остроты зрения при старении.
То положение, что различительная чувствительность глаза лимитируется шумом нервной системы, имеет далеко идущие выводы. Из него следует, что старое представление о пороговых интенсивностях, которых должны достичь стимулы, прежде чем они вызовут какой-либо ответ нервной системы, — неверно. Сейчас мы считаем, что каждый стимул оказывает воздействие на нервную систему, но он воспринимается как внешний сигнал только тогда, когда вызываемые им изменения нервной активности превосходят обычный уровень шума. Пример этого можно видеть на рис. 6, 10. На нем изображено пятно света, являющееся исходным фоном (I), к которому прибавляется различимый свет (ΔI). Эти две интенсивности света приводят к появлению нервных импульсов, частота которых подчиняется статистическим закономерностям.
Рис. 6, 10. Здесь мы пытаемся показать, что действие статистических закономерностей в мозгу возникает вследствие случайной активности нервной системы. Когда сигнал (ΔI) выделяется из более слабого фона (I), частота импульсов не всегда является иной, но имеет такое распределение, как это показано на графике. Таким образом, можно увидеть «свет» на фоне «шума» или не замечать его, если частота импульсов ниже средней. Различие между сигналом и шумом должно быть достаточным для того, чтобы мозг расценил нервную активность как сигнал.
Проблема, возникающая перед мозгом, состоит в том, чтобы «решить», когда увеличение числа импульсов является просто случайным, а когда оно возникает вследствие увеличения интенсивности светового сигнала. Если бы мозг принимал любое увеличение числа импульсов по отношению к средней активности за объективный сигнал, тогда мы «видели» бы вспышки света, отсутствующие в действительности, по крайней мере, в половине случаев. Таким образом, мы приходим к мысли, что не обходимы некоторые значимые различия, чтобы возникшая нервная активность оценивалась как результат воздействия сигнала. Наименьшее различие освещенности (ΔI), которое мы можем видеть, определяется не просто чувствительностью рецепторов сетчатки, но также и различием в частоте нервных импульсов, необходимым для того, чтобы воспринять его как сигнал.
Иногда мы видим вспышки, которых на самом деле нет. По-видимому, они появляются вследствие шума, переходящего требуемый уровень значимости, вследствие готовности к восприятию сигнала, но это случается но часто.
Определение уровня, выше которого активность принимается за ответ на реальное воздействие, и используется для оценки надежности данной чувствительной системы. Существуют доказательства того, что этот уровень может колебаться и зависит от нашей «установки». Когда мы особенно осторожны, требуется большая информация и чувствительность снижается.
То, что сказано выше по поводу восприятия интенсивностей света, применимо к нервной системе в целом. Bee это справедливо не только для различения интенсивностей света, но также и в отношении абсолютного порога различения света в темноте. Абсолютный порог также определяется наименьшим сигналом, который может быть надежно выделен из случайного шума зрительной системы, существующего в мозгу и при отсутствии воздействия света на глаз.
Более 800 000 книг и аудиокниг! 📚
Получи 2 месяца Литрес Подписки в подарок и наслаждайся неограниченным чтением
ПОЛУЧИТЬ ПОДАРОК