СО2 + Н2О + свет = жизнь
Солнечный свет, пронизывающий толщу моря, приводит в действие, пожалуй, самый удивительный и совершенный механизм в природе — механизм преобразования неорганических веществ (углекислого газа и воды) в органическое соединение — углевод. Этот процесс, при котором растение, используя свет в качестве энергии для осуществления синтеза различных веществ в клетке, вступает в окислительно-восстановительную реакцию с CO2 и Н2O, был назван фотосинтезом.
Суммарный конечный результат его можно представить следующим уравнением:
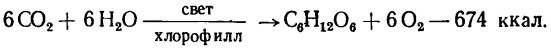
В результате этой реакции возникают углеводы, обеспечивающие рост растений, а в окружающую среду выделяется свободный кислород.
В морях и океанах обитают многочисленные водоросли, способные к фотосинтезу. Некоторые, наиболее крупные из них, ведут оседлый образ жизни в прибрежной полосе океана. Но основную массу морских растений составляют не эти донные водоросли. Главный поставщик пищи в море — планктонные водоросли, мельчайшие одноклеточные (или колониальные) растения, проводящие всю свою жизнь в толще воды. Их морфологическое строение и удельный вес, близкий к удельному весу морской воды, позволяют им «парить» в море. Нуждаясь в свете, они обитают только в тонком поверхностном слое, до глубины 100, реже 200 м, где достаточно лучистой энергии для поддержания фотосинтеза. Образную картину процесса фотосинтеза, рисующую его исключительную важность для всего живого, дал великий русский ученый К. А. Тимирязев: «Когда-то, где-то на землю упал луч Солнца, но он упал не на бесплодную почву, он упал на зеленую былинку пшеничного ростка, или, лучше сказать, на хлорофилловое зерно. Ударяясь о него, он потух, перестал быть светом, но не исчез. Он только затратился на внутреннюю работу, он рассек, разорвал связь между частицами углерода и кислорода, соединенными в углекислоте. Освобожденный углерод, соединяясь с водой, образовал крахмал. Этот крахмал, превратясь в растворимый сахар, после долгих странствий по растению отложился, наконец, в зерне в виде крахмала, или в виде клейковины. В той или другой форме он вошел в состав хлеба, который послужил нам пищей. Он преобразился в наши мускулы, в наши нервы»[33].
Естественно, что в процессе дальнейших исследований механизм фотосинтеза был уточнен. Так, было доказано, что солнечная энергия расходуется на разложение не углекислоты, а воды.
Что же происходит под воздействием света в водах Мирового океана?
Солнечная энергия, проникая в море, так же как и на суше, поглощается и используется растениями. За счет этой энергии происходит образование восстановленных органических соединений из окисленных неорганических. Энергия, накопленная в этих соединениях, покрывает все энергетические затраты организмов, населяющих море, во время их движения, размножения, биохимического обмена и т. п. Передача энергии, усвоенной растениями, происходит и когда одни организмы питаются другими.
Первоисточником пищи всех животных служат органические соединения, накопившиеся в теле водорослей в процессе фотосинтеза. Это накопление органических соединений, приводящее к росту водорослей, называют первичной продукцией океана.
В ходе процесса фотосинтеза в клетке происходит фотохимическое расщепление молекулы воды на ионы гидроксила и водорода. Водород присоединяется к углекислоте и образует углеводы. Затем клетка извлекает из воды азот, фосфор, калий, кальций, магний, серу и др. Образуются белки и другие биохимические соединения. Интересно отметить, что все процессы, кроме фотодиссоциации воды, темновые, т. е. не требуют дополнительного поглощения световой энергии. Продолжительность темновых реакций во много раз больше, чем световых. Суммарная скорость процесса первичной продукции в условиях достаточного снабжения биогенными элементами и хорошей освещенности в конечном счете определяется темновыми биохимическими реакциями.
Если света мало, фотохимические (световые) реакции оказываются определяющими, лимитирующими общую скорость фотосинтеза.
Поглощение солнечной энергии растениями осуществляется с помощью пигментов, т. е. каталитически активных окрашенных соединений, содержащихся в водорослях. Каждый из пигментов поглощает свет в определенных, характерных для него участках спектра.
Основным из них является хлорофилл a, который есть у всех без исключения фотосинтезирующих растений. Молекулы хлорофилла — отличные «ловушки» для солнечного света. Но главная их функция — фотодиссоциация молекулы воды. Наиболее ярко выраженные максимумы поглощения хлорофилла a находятся в участках спектра 420–430 и 670–680 нм. Однако эти области излучения значительно поглощаются морокой водой, и поэтому у водорослей имеется сложная пигментная система, восприимчивая к излучению и в других участках видимого спектра. Дополнительные пигменты поглощают энергию в участках спектра, недоступных хлорофиллу а, и передают ему эту энергию. От набора пигментов данной водоросли зависит ее окраска. Окраска водорослей — наследственный признак и способствует их расселению на определенных глубинах.
Важным свойством пигментной системы является ее способность к хроматической адаптации, т. е. способность быстро изменяться и приходить в соответствие с суточными и вертикальными изменениями спектрального состава проникающего в воду света.
В процессе фотосинтеза морского фитопланктона используется лишь очень незначительная доля световой энергии, падающей на поверхность Мирового океана, — в среднем 0,04 % от проникшей в воду. В наиболее продуктивных районах моря эффективность фотосинтеза достигает примерно 0,35 %, а в бедных жизнью она составляет всего 0,02 %. Остальная солнечная радиация поглощается водной толщей.
Как мы знаем, освещенность с глубиной убывает по показательному закону. Тот же характер носит ослабление с глубиной интенсивности фотосинтеза. Происходит это до тех горизонтов, где световые реакции фотосинтеза уже начинают лимитировать его скорость. В подповерхностных же слоях моря, наоборот, избыток света подавляет фотосинтетический процесс. Исключение составляют лишь приполярные районы, где на поверхность моря падает количество световой энергии, которого недостаточно для подавления фотосинтеза.
Оптимальное для фотосинтеза значение световой энергии для различных биогеографических областей неодинаково: для тропической зоны это 60–85 кал/см2, для умеренных широт — 15–20 кал/см2 в день. Минимальная интенсивность освещения, при которой вообще возможен фотосинтез, очень невысока. Многие виды глубоководных морских водорослей развиваются в условиях, где освещенность составляет 10-5—10-7 от количества солнечного света, падающего на морскую поверхность, что по интенсивности близко к освещенности, создаваемой лунным светом.
Таким образом, интенсивность световой энергии на различных глубинах моря, ее суточные и сезонные изменения имеют большое значение для первичной продукции вод океанов и морей.
Важной величиной, характеризующей эффективность превращения световой энергии в химическую, является квантовый выход фотосинтеза ?, который можно оценить числом молекул синтезированного углерода при поглощении одного кванта света, т. е.
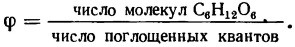
Квантовый выход фотосинтеза меньше единицы, поэтому удобнее пользоваться обратной величиной (1/? = n), называемой квантовым расходом и показывающей, сколько квантов света затрачено на синтезирование одной молекулы углерода C6H12O6— гексозы.
Из графика на рис. 66 видно, что квантовый выход фотосинтеза неодинаков на всем участке видимого спектра. В наиболее благоприятных условиях он не превышает 0,12, т. е. необходимо примерно 8 квантов света (n = 1/0,12) для синтезирования одной молекулы гексозы. Но величина эта непостоянна и колеблется в довольно широких пределах, определяемых целым комплексом условий, в которых протекает процесс фотосинтеза.
При менее благоприятных условиях, чем во время проведения опытов (недостаток элементов питания, света и т. п.), квантовый выход падает.
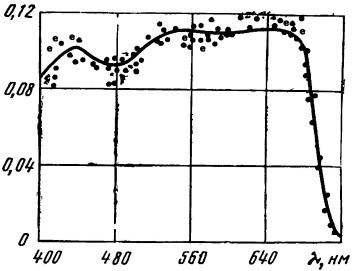
Рис. 66. Спектральная зависимость квантового выхода фотосинтеза
Вообще говоря, выяснить зависимость первичной продукции в океанах от условий подводной освещенности — задача исключительной трудности.
Есть еще очень много неясного в сложнейшем процессе воздействия лучистой энергии на морской фитопланктон. Какова реакция живой клетки водорослей на излучение различных длин волн, как происходит адаптация пигментной системы в зависимости от условий освещения и т. п. — все эти вопросы относятся главным образом к сфере деятельности ученых, изучающих физиологию растений. Гидрооптике как одной из отраслей физической океанологии в комплексе этих проблем отводится несколько особая роль.
В исследовании световой энергии, распространяющейся в море, и ее влияния на фотосинтез морских водорослей есть один весьма важный аспект.
Дело в том, что производство гидрооптических измерений, сопутствующее биологическим исследованиям, ведется практически на всей обширной акватории Мирового океана учеными разных стран.
В своих исследованиях они применяют измерители энергии самых разнообразных конструкций и используют различные методы для расчетов как суммарной энергии, так и ее спектрального состава. В результате полученные данные практически мало сопоставимы друг с другом, а это в свою очередь мешает ученым получить реальную картину зависимости процесса первичной продукции от условий освещенности в масштабах всего океана.
Необходимость унифицировать проведение световых измерений в море привела к созданию по инициативе ЮНЕСКО, Научного комитета по океанографическим исследованиям (СКОР) и Международной ассоциации физической океанографии (МАФО) Рабочей группы по фотосинтетической радиации в море. В ее состав вошли специалисты по первичной продукции Г. Джиттс (Австралия), Е. Стимен-Нильсен (Дания), Ю. Сайджо (Япония), Дж. Стил (Шотландия) и по оптике моря Н. Ерлов (Швеция), А. А. Иванов (Франция), Ю. Е. Очаковский (СССР) и Дж. Тайлер (США).
Эта группа должна решить два основных вопроса: во-первых, какие измерения световой энергии в море надо проводить, чтобы установить закономерности, существующие между процессом фотосинтеза и светом в море, и, во-вторых, какая аппаратура способна обеспечить нужную точность и сопоставимость измерений.
В октябре 1964 г. в Москве состоялось первое совещание. Естественно, рассматриваемые Вопросы вызвали довольно бурную дискуссию.
Председатель группы Дж. Тайлер высказал предположение, что в основу искомого прибора должен быть положен приемник излучения, обладающий спектральной чувствительностью, такой же как спектральная чувствительность живой клетки фитопланктона.
Его точка зрения была принята буквально в штыки биологами. Они заявили, что создание прибора на этом принципе невозможно по многим причинам. Первая и основная: видовое разнообразие планктонных организмов столь велико и их спектральная чувствительность к излучению настолько различна, что подобрать какую-либо одну, осредненную кривую, удовлетворительную для всех случаев, просто не реально. Кроме того, реакция планктона на свет не остается постоянной, так как он обладает способностью к световой и хроматической адаптации, причем надо еще учитывать инерционный характер этих приспособительных процессов.
После длительного обсуждения было принято решение, обязывающее гидрооптиков разработать конструкцию такого прибора, который позволял бы с точностью до 10 % измерять суммарную лучистую энергию, распространяющуюся от поверхности моря к его глубинам в диапазоне 350–700 нм. Такой измеритель должен быть прост в обращении и доступен в массовых исследованиях. Кроме датчика, погружаемого в море, прибор необходимо снабдить аналогичным приемником излучения для измерения радиации в палубных инкубаторах, где имитируются условия освещения на различных глубинах.
Кроме того, прибор, который предстояло создать, должен обладать способностью суммировать энергию за время экспозиции склянок с пробами. Задача не из легких. Но, пожалуй, самое трудное — это добиться, чтобы приемник излучения прибора был неселективным в пределах 350–700 нм и не реагировал на излучение вне этого диапазона, т. е., чтобы он измерял лучистую энергию, проникающую в море, именно в данном участке спектра.
Прежде чем окончательно решить вопрос о создании нового прибора, надо было проверить, нельзя ли использовать уже существующую аппаратуру для непосредственных измерений энергии, а также один из косвенных методов расчета ее значений — оптическую классификацию вод, предложенную Ерловым.
Действительно, если известна закономерность, с которой в данных водах происходит ослабление лучистой энергии в различных участках спектра, то достаточно измерить энергию, падающую на поверхность моря, и ее ослабление в воде в одном узком участке спектра, с тем чтобы, используя кривые Ерлова, рассчитать суммарную энергию на требуемой глубине.
Для этих целей надо установить на палубе пиранометр стандартного типа, а для измерения показателя вертикального ослабления воспользоваться примитивным фотометром с селеновым фотоэлементом, перекрытом узкополосным цветным светофильтром.
В лаборатории гидрооптики ИОАН были проведены методические исследования, которые показали, что в прозрачных водах открытого моря метод оптической классификации вполне применим, но его точность несколько снижается в более мутных прибрежных водах.
И вот прошло четыре года. В мае 1968 г. гидрооптики, члены Рабочей группы, встретились в Сан-Диего (США), чтобы сопоставить различные приборы для измерения фотосинтетически активной радиации (ФАР) в море. Одновременно австралийский гидробиолог Джиттс измерял первичную продукцию, использовав палубный инкубатор своей конструкции.
Испытания проводились в Калифорнийском заливе; штилевая погода, отсутствие облачности и большие высоты Солнца создавали хорошие условия для работы.
Измерения велись различными по конструкции и принципу действия приборами.
Американские ученые Тайлер и Смит измеряли и спектральное распределение энергии на различных глубинах с помощью погружаемого в море спектрорадиометра. Этот прибор позволял получать спектральные кривые большой точности, так как его разрешающая способность была всего 5 ям. Аналогичный прибор использовали и французские гидрооптики Иванов и Боэр. Кроме того, они испытали подводный пиранометр повышенной чувствительности и измеритель облученности в узком участке спектра. Советские специалисты Очаковский и Сусляев использовали подводный пиранометр, измеритель облученности в узком участке спектра и прибор ВАРИПО.
Особый интерес вызвал измеритель Ерлова, названный им квантометром. Приемником здесь служит селеновый фотоэлемент, различные участки поверхности которого перекрыты разными цветными светофильтрами. Такая конструкция, по мнению Ерлова, делает приемник излучения неселективным в диапазоне ФАР. К сожалению Ерлов не смог принять участие в испытаниях, и с этим прибором работали его коллеги К. Ньюгард и Г. Кулленберг.
Сейчас, когда полученные материалы еще находятся в стадии обработки, трудно судить о достигнутых результатах. Но очевидно, что это только один из первых шагов в поисках оптимального варианта прибора для световых измерений, которые так необходимы морским биологам.
Интересную аппаратуру для изучения фотосинтетически активной радиации сконструировал польский ученый Ежи Дера. Его установка, плавая в виде поплавка, суммирует количество энергии, падающей на поверхность моря в течение всего дня.
Надо помнить, что проблема «свет и фотосинтез» остается одной из важнейших проблем как оптики моря, так и морокой биологии. Океан — продовольственная кладовая будущего, и ее изобилие в первую очередь определяется первичной продукцией.
Недавно советские ученые О. И. Кобленц-Мишке, В. В. Волковинский и Ю. Г. Кабанова примерно оценили первичную продукцию Мирового океана. Оказалось, что в год морские водоросли переводят из неорганического состояния в органическое около 15–20 млрд. т углерода. Другими славами, водоросли за один год накапливают 600–800 млрд. т сырой биомассы.
При этом надо учесть, что биологические ресурсы моря пока не испытывают регулирующего влияния человека. В дальнейшем, когда люди научатся культивировать моря и океаны, их продуктивность, несомненно, еще больше возрастет.
Более 800 000 книг и аудиокниг! 📚
Получи 2 месяца Литрес Подписки в подарок и наслаждайся неограниченным чтением
ПОЛУЧИТЬ ПОДАРОК