Популяционно-генетические исследования манси
Г. М. Давыдова
В последние годы Институт этнографии АН СССР проводил антропологические исследования среди мансийского населения. Общая численность манси составляет около 5 тыс. человек. Подавляющее большинство манси живет в двух районах Тюменской области: в Верезовском (примерно 3 тыс. человек, включая метисов) и Кондинском (1,5 тыс. человек) и незначительное число — в Свердловской области (чуть больше ста человек).
Была обследована группа манси Березовского района, расселенная по Северной Сосьве и ее притоку Липину, по развернутой антропологической программе, включающей и серологическую часть. Определялись следующие системы крови: ABO, MN, Р, Lewis, Резус, Kell-Cellano. У небольшой части исследованных определения резус-системы производилось с помощью пяти сывороток: анти-D, анти-С, анти-Е, анти-с, антн-е (образцы отсылались в лабораторию М. А. Умновой в Центральный Институт гематологии и переливания крови), остальные определения производились на месте в день получения образцов или па следующий по обычным методикам.
Численность манси Березовского р-на не изменилась на протяжении последних полутора столетии.
Большинство браков заключаются внутри мансийской популяции, однако смешанные браки — далеко не редкость. Число смешанных браков меньше в верховьях Северной Сосьвы (в поселках Няксимвольского сельсовета) и по Ляпипу — соответственно 13,8 и 11,9% (данные похозяйственных книг). В поселках среднего и нижнего течения Северной Сосьвы частота сметанных браков выше и составляет более четверти всех браков (27,U% в Сосьвинском сельсовете и 32,2% в Банзетурском).
Таким образом, смешение манси с соседями получает большой размах. Наиболее часты браки сосьвинских манси с русскими — 6,92 %, коми-зырянами — 6,22 %, хантами — 4,38 %, ненцами — 2,53 % > эвенками — 0,23%, немцами — 0,23%, калмыками — 0,23 %, украинцами — 0,23 %, татарами — 0,23 %. Всего смешанные браки у сосьвинских манси составляют 20,20%.
Мы исследовали только лиц чисто мансийского происхождения, насколько это можно было установить.
Сосьвинских манси можно разделить на три территориальные группы, в известной мере обособленные друг от друга: ляпинскую, живущую по Ляпину, верхнесосьвинскую, живущую в верховьях Сосьвы от У-Маньи до Нильдино, и среднесосьвинскую, куда входят жители поселков от пос. Сосьва до Ванзетура. Названные группы мапси несколько различаются и по особенностям языка.
Самая обособленная среди этих групп — ляпинская, здесь 94% всех браков между манси заключалось внутри группы. На среднем течении Северной Сосьвы внутри групны заключалось 89 % браков. Приток в этот район идет в основном с Ляпина— в 8,2% браков один из супругов происходит с Ляпина. Верхнесосьвннская популяция больше, чем две другие, включает в себя лиц, происходящих из других мест, в 18% браков один из супругов — уроженец соседней местности: в 7% —из Ивдель-ского района Свердловской области, в 5,5% —с Ляпина и в таком же количестве со среднего и нижнего течения Северной СоСЬВЫ.
Суммарная гематологическая характеристика березовских манси и их отдельных групп приводится в табл. 1.
Система АВ0. Березовские манси (суммарная группа) в сравнении с другими народами Сибири и Поволжья характеризуются довольно высокой частотой гена г (0,629) и небольшой частотой гена р. Приближаются к ним по частотам генов системы ABО Некоторые группы ненцев, якуты (суммарно) и эскимосы. Впрочем, манси можно объединять в одну суммарную группу по системе АВ0 лишь условно, так как отдельные группы значительно различаются между собой. Наиболее резко уклоняется от других
Фенотипы и гены Сосьвинские НСШСИ (суммарно) Ляпинскаягруппа Среднесось-винскангруппа Верхнееосьвнн ская группа N частота N частота N частота N частота 0 эмп. 426 0,3967 155 0,4968 166 0,3675 105 0,2952 теор. 0,4057 0,5092 0,3670 0,3112 А эмп. 0,2535 0,2258 0,2711 0,2667 теор. 0,2430 0,2119 0,2717 0,2323 В эмп. 0,2858 0,2516 0,2831 0,3810 теор. 0,2857 0,2380 0,2836 0,3622 AB эмп. 0,0540 0,0258 0,0783 0,0571 теор. 0,0656 0,0409 0,0777 0,0943 12 1,2200 1,1700 0,0013 1,7800 эмп. 0,6290 0,7048 0,6062 0,5433 г теор. 0,6370 0,7136 0,6060 0,5578 р эмп. 0,1680 0,1349 0,1934 0,1777 теор. 0,1680 0,1356 0,1932 0,1794 q эмп. 0,1940 0,1500 0,2009 0,2604 теор. 0,1950 0,1508 0,2008 0,2628 М эмп. 426 0,2019 155 0,1806 166 0,2048 105 0,2286 теор. 0,2095 0,1840 0,2041 N эмп. 0,2864 0,3226 0,3012 0,2095 теор. 0,2940 0,3260 0,3005 MN эмп. 0,5117 0,4968 0,4940 0,5619 теор. 0,4965 0,4900 0,4954 т 0,4578 0,4290 0,4518 0,5100 D- 388 0,0284 138 0,0290 144 0,0417 106 0,0094 d 0,1682 0,1702 0,2042 0,0970 Kell* (KK+Kk) 228 0,0219 102 0,0099 126 0,0317 — — К 0,0110 0,0050 0,0160 — Р— 421 0,2880 150 0,2267 166 0,3433 105 о.зззз Р 0,5470 0,4762 0,5859 0,5773 Le а—Ь+ 422 0,6256 149 0,6174 164 0,6158 109 0,6514 Le а--В— 0,0498 0,0431 0,0610 0,0459 Le а—b— 0,3246 0,3423 0,3232 0,3028 Le“ 0,2122 0,2076 0,2470 0,2142ляпинская группа, отличаясь статистически реально от верхне-сосьвинской (х2 = 6,68 0,95Р = 0,0975) и почти реальпо от среднесосьвпнской (х2 = 5,89, при Р=0,95х2 должен быть равен 5,99).
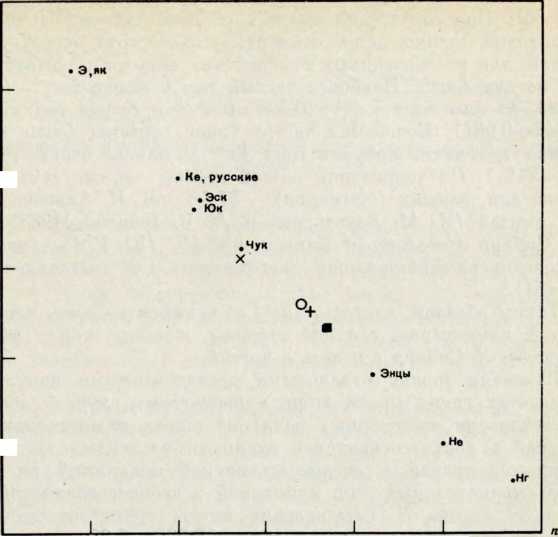
Ляппнские манси, наиболее обособленная популяция среди трех нами исследованных, выделяются среди других березовских манси высокой частотой гена г и небольшими частотами генов р и q. Частота гена q у ляпинских манси примерно равна средней частоте его у русских, у некоторых финских групп на европейской территории страны и ниже, чем у поволжских народов (удмуртов, мари, чувашей, татар), однако выше, чем у народов Северной Сибири. Эти особенности распределения частот генов г, р, q ставят ляпинских манси в особое положение среди всех исследованных народов Сибири и Поволжья (рис. 1).
Рис. 1. Частоты генов г, р, q у манси и в сравниваемых группах
Ал — алтайцы, Бу — буряты, На — карелы, Не — кеты, Из — коми-зыряне,
Ло — лопари, Ма — марийцы, Мм — мордва-мокша. Л/а — мордва-эрзя, Нг — нганасаны, Неа — ненцы архангельские, Та — татары казанские, Ув — удмурты, Ф — финны Ленинградской области, Чу — чуваши, Чу к — чукчи, Э — эвены, Эв — эвенки, Эск — эскимосы, Юк — юкагиры, Як — якуты суммарно.
1 — манси суммарно, г — манси ляпинские, з — манси средпесосьвинские, 4 — манси верхнесосьвинские
В верхнесосьвинской группе манси высока частота гена Я (0,263); подобные частоты встречаются редко, в частности у бурят (0,265), у архангельских ненцев (0,282), у марийцев (0,246).
В общем частоты гена q у березовских манси довольно высокие, даже в группе с наименьшими значениями q.
Сибирские народы довольно четко делятся на две группы По частоте гена q. Как отмечала И. М. Золотарева (И. М. Золотарева, А. Г. Башлай, 1968) у палеоазиатских и других народов Северной Сибири частота гена q низка; у эвенков — 0,078 (Ю. Г. Рычков, 1965); у эвенов — 0,134 (И. М. Золотарева, 1964; М. Г. Левин, 1959); у чукчей — 0,095 (М. Г. Левин, 1958; И. М. Золотарева, 1964); у юкагиров — 0,064 (И. М. Золотарева, 1964); у нганасан — 0,133 (И. М. Золотарева, 1964). У южносп-бирских народов (или народов, связанных по происхождению с Южной Сибирью) частота гена q высокая: у якутов — 0,209 (И. М. Золотарева, А. Г. Башлай, 1968), в Забайкалье у бурят 0,265 (В. В. Бунак, 1963), у алтайцев суммарно — 0,202, у ал-тай-кижи — 0,245 (Рычков, 1965). По нашим данным, у алтай-кнжи q равно 0,277 (УУ = 241) и у теленгетов — 0,137 (N=81), Повышенная частота гена q наблюдается также у кетов — 0,183 (И. И. Гохман, 1963) и у архангельских ненцев — 0,282 (В. В. Бунак, 1969), т. е. у народов, в составе которых прослеживаются южные элементы.
Другая большая область расселения народов с высокой частотой гена q — север европейской территории нашей страны и Поволжье — серологический «тип, сложившийся на месте в далеком прошлом» (В. В. Бунак, 1969, стр. 32).
Манси, одна из древних групп Западной Сибири, отличается от палеоазиатских и других народов Северной Сибири значительно более высоким содержанием гена q и примыкают по этому признаку к народам Поволжья, с которыми они сближаются также и по антропологическому типу.
Однако возможно, что некоторые группы манси впитали в себя примесь иного этноса, монголоидного по своему физическому типу, видимо южного, имевшего большую частоту гена q. Это предположение касается в основном верхнесосьвпнскои группы, в которой, как уже указывалось, отмечена самая высокая у манси частота гена q; в ней же наблюдается некоторый сдвиг в сторону большей уплощенности лица и носа, более темной пигментации волос и глаз, меньшего роста бороды.
Частота гена q у ляпинских манси, несколько меньшая, чем у сосьвинских, возможно, свидетельствует о наличии палеоазиатских элементов в этой группе.
В конце 30-х годов Н. П. Чебоксаров и Т. А. Трофимова определяли группы крови у ивдельских и пелымских манси. Распределение частот генов г, р, q у них сильно отличается от того, что мы нашли у березовских манси. В ивдельской группе очень высокая частота гена г (0,817) и низкие частоты р (0,108) и q (0,084), т. е. эта популяция манси отличается от остальных в том же направлении, что и ляпинская, но намного сильнее. У пелымских манси частота гена г — 0,588, р — 0,264 и q - 0,148, т. е. частота гена q невысокая, а р — выше, чем в березовских популяциях. Очень близкие частоты генов rpq (0,589, 0,256 п 0,155 соответственно) тогда же были обнаружены у ваховскпх хантов (Дебец, 1941).
Система MN. Частота гена m у березовских манси составляет немного менее 50% (0,458), колебания его в отдельных группах (от 0,429 в ляпинской до 0,510 в верхнесосьвинской) и не достигает статистически достоверных величин.
У сибирских народов частота гена m колеблется в очень широких границах — от 0,260 у нганасан до 0,721 у якутов. При этом выделяются два типа частот гена m: у одних народов — ненцев, нгапасан — ген п преобладает над m, у других — тп преобладает над п либо резко, как у якутов, эвенов, либо незначительно, как у юкагиров, кетов, чукчей, эскимосов (И. М. Золотарева, 1964; И. М. Золотарева, А. Г. Башлай, 1968). Превышение частоты гена m над п характерно и для алтайских народов; у алтай-кпжп частота гена m составляет 0,670 (N=238), у теленгетов — 0,648 (W=81).
Таким образом, манси занимают по частоте гена m довольно своеобразное место среди сибирских народов: между энцами, ненцами и нганасанами, с одной стороны (причем на довольном удалении от всех их), и чукчами и эскимосами, с другой (также в среднем на значительном удалении), и очень сильно отдалены от всех других народов Сибири (рис. 2). Можно предположить, что такие частоты гена m у манси определяются наличием у них палеоазиатского элемента, может быть, того же, что у ненцев и нганасан.
Отмечается несколько более высокая частота гена m в верхнесосьвинской популяции (0,510), что наряду с высокой частотой гена q делает вероятным предположение о наличии южносибирской монголоидной примеси в этой группе.
Среднесосьвннская группа по частотам генов систем АВ0 и MN занимает промежуточное положение между ляпинской и верхнесосьвинской (см. рис. 2).
Система Rh. Резус-принадлежность определялась в 388 случаях с помощью одной сыворотки (анти-D) и в 72 — с помощью пяти сывороток. Частота резус-отрицательных лиц в мансийских популяциях составляет 1–4%. Наименьшая частота резус-отрицательных лиц (0,94%) найдена в верхпесосьвннской группе, это частота такого порядка, которая была обнаружена у якутов (И. М. Золотарева, 1964; В. П. Алексеев и др., 1968), бурят (В. В. Бунак, 1963). В двух других популяциях манси резус-отрицательных лиц встречено больше (3–4%).
Табл. 2 показывает, что у манси Березовского р-на встречаются только три хромосомы: cDe, CDe и cDE. В этой выборке практически нет хромосомы cde. Распространение частот генов па земном шаре по литературным материалам рассматривает Ю. Д. Беневоленская (табл. 3).
Манси по резус-системе не занимают промежуточного положения между европейским и азиатским комплексами. У них частоты /?/г-хромосом найдены в таком сочетании, которое не встречено больше ни в одной группе: хромосома cDE найдена
Фенотип N Частота в °/o Хромосома Частота в % эмпирическая * * эмп. теор. CcDee 13 18,05 13,60 cDe 16,67 CcDEe 20 27,78 30,38 CDe 31,18 CCDee 7 9,72 10,66 cDE 48,59 ccUEE 17 23,61 21,65 — — ccDEe 12 16,67 19,38 — — ccDee 2 2,78 4,34 — — ccddEe 1 1,39 — — — Всего 72 * Материалы Г. М. Давыдовой и М. А. Умновои.** Вычисления сделаны по способу Моранта (Mourant, 1954). Таблица 3. Пределы вариаций генов резусных комплексов Хромосома Европейский Африканский Азиатский Американский Австралийский Мела 110-пндонезпй-ский Л1 (CDe) R2 (сПЕ) Д° (сDe) R* (CDE) г (cde) 0,40–0,500,10–0,180,01–0,040,00–0,010,35–0,40 0,03–0,190,01–0,180,60–0,900,000,06–0,24 0,40–0,550,30–0,400,00–0,150,00–0,020,00–0,09 0,33–0,600,30–0,600,00–0,070,00–0,080,00–0,09 0,55–0,700,12–0,380,02–0,160,00–0,080,00 0,78–0,97 0,02–0,15 0,00 -0,07 0,00–0,02 0,00в количестве, характерном для индейских популяций, частота хромосомы CDe также близка к частотам американского комплекса, частота cDe немного превышает частоту этой хромосомы в Азии. Частоты ДЛ-хромосом в саянских группах пе имеют никакого сходства с распределением их в мансийской выборке.
Как сказано выше, у основной массы манси ЯА-принадлежность определялась с помощью одной сыворотки — анти-?. Наименьший процент резус-отрицательных лиц, найденный в верхнесосьвинской группе, сочетается в ней с большой частотой гена g и с более высокой, чем в двух других, частотой гена т, что отличает эту мансийскую популяцию от остальных. Средний процент резус-отрицательных лиц у манси (2,9%) выше, чем у бурят, якутов, и совпадает с таковым у алтай-кижп Онгудайского р-на (2,95%, iV=241).
Система Р. Частота гена Р (Р) составляет у березовских манси в среднем 45,3%, в ляпинской популяции процент этого гена равен 52,4, в верхнесосьвинской — 41,4.
0,700
0,600 —
0,500
0,400
0,300 -
0,200 0,300 0,400 0.500 0,600 0,700
Рис. 2. Частоты генов т и п у манси и в сравниваемых группах Обозначения те же, что на рис. 1
Частоты гена Pi у европейцев колеблются от 46% У австрийцев до 57,1% у датчан (A. Mourant, 1954); у русских она составляет 48,5% (М. А. Умнова и др., 1964). У азиатских народов процент гена Рi более низкий: у китайцев — 15–18% (A. Mourant, 1954), у якутов — 30,1 (И. М. Золотарева и А. Г. Башлай, 1968), у тофаларов — 26,7%, тоджинцев — 17,9%, тувинцев — 38,0% (Ю. Г. Рычков п др., 1969).
Геп Pi в ляпинской популяции встречается с частотой, средней для европейского населения; в двух других группах манси частота его ниже, чем у европейцев и выше, чем у азиатских народов. У живущих на Ляпине коми-зырян геп Pi (N=52) встречен в 50% случаев. Думается, что более низкие частоты гена Pi в двух сосьвинских популяциях манси говорят о влиянии южносибирских монголоидных популяций на эти группы.
Система Kell-Cellano определялась с помощью сыворотки анти-/!Г в двух группах манси: ляпинской и сосьвинской, частота гена К найдена равной 0,5 и 1,6% соответственно.
У якутов (И. М. Золотарева, А. Г. Башлай, 1968) частота гена К равна 2,07%, у русских — 4% (М. А. Умпова и др., 1964: А. Г. Багплан, 1968), у коми-зырян, живущих на Сосьве, — 11,8% (N=36). При отсутствии данных об этой системе со смежных территорий можно лишь отметить, что частота гена К у манен низкая, для исследованных европейских народов не характерная.
Система Lewis. Наиболее частый тип у манси Le“-6* (0,616–0,651), за ним идет Le“ ь_ (0,303–0,323) и самый редкий Le°+b (0,043–0,061). Колебания частот типов системы Lewis в популяциях певелики, процент гена Le“ у манси очень стабилен (21–25%). На территории нашей страны частота гена Le“ из вес.тпа для русских (суммарно) —37,1% (JT. К. Аржелас, 1965), для якутов (И. М. Золотарева и А. Г. Башлай, 1968) — 16,5, для Сибири к востоку от Енисея — 13,4% (10. Г. Рычков, 1965V У сосьвпнскпх комп-зырян частота гена Le° составляет 28,0% (7V=51).
Таким образом, частота гена Le“ у манси меньше, чем у русских и комп-зырян, с одной сторопы, и выше, чем у якутов и в Восточной Сибири в целом, с другой.
Различия между отдельными исследованными популяциями в частотах групп крови лишь в нескольких случаях достигают статистически достоверных величин: такова разница между ля пинской и верхнесосьвинской группами по гепам rpq, различия верхпесосьвипской и среднесосьвппской популяций по частоте резус-отрицательпых лиц, ляпинской и средпесосьвинскон групп по частоте гепа Р. Расхождения между группами в немпогнх случаях вероятны па уровне между 0,90 и 0,95 — это разиипьг в частотах генов rpq в ляпппской и среднесосьвинской группах п частотах гепа Р в ляпинской и верхпесосьвипской. Остальные различия между популяциями по группам крови далеки от ста тпетичеекп реальных.
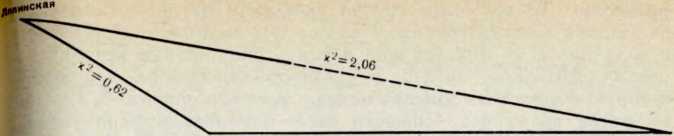
Сопоставлеппе популяций друг с другом по всем группам крови не дает статистически существенных различий между нпмп, однако показывает, какие популяции оказываются ближе друг к другу и какие сильнее удалены одна от другой. Для та кого сравпеппя был примепен прием, использованный Кнуесмп ном (I. Schwidetzki и др., 1962). Выяснилось, что верхнесосьвип ская популяппя по сумме систем крови удалепа от двух другп более всего, папболыпее расстояние отделяет ее от ляпппской группы; мепыпе удалены друг от друга ляпинская п среднесось впнекая популяции (рис. 3).
Близость ляпинской и среднесосьвппской групп находит свое объяснение в том, что брачные круги этих двух групп Пересе каются: жители среднего течения Сосьвы довольно часто вступают в брак с жптелямп Ляпппа.
В верховьях Северной Сосьвы отмечены брачные связтт с мапси Ивдельского р-па Свердловской области. Можпо было бы предположить, что пмеппо этот факт нашел отражепие в большей удаленности верхнесосьвинской группы от двух остальных. Однако частоты генов г, р и q у пвдельекпх манси (материалы
Среднесосьвинская х “ 1,29 Верхмесосьаииская
рис. 3. Удаленность мансийских популяций друг от друга но сумме генов крови
H. Н. Чебоксарова и Т. А. Трофимовой, указанные выше) таковы, что если бы здесь сказывалось их влияние, то оно сблизило бы верхнесосьвпнских манси с остальными. (Правда, к материалам по крови, полученным в 30-е годы, надо подходить с некоторой осторожностью вследствие несовершенства методики определения групп крови в те годы (К. Штерн, 1965), однако возможные ошибки вряд ли существенно меняют общую картину.)
Три исследованные группы манси, как указано выше, различны по величине брачного круга. Он более узок в Ляпинской группе: 94% всех браков заключается здесь внутри группы. Наиболее широкий круг брачных связей отмечен в верхнесось-винской группе. Можно думать, что если из поколения в поколение браки заключаются внутри какой-то группы, то через некоторое время все члены этой группы неизбежно окажутся в родстве между собой и группа достигнет какого-то уровня инбридинга, даже при отсутствии браков между близкими родственниками. Если в некоторой части браков один пз супругов происходит из другой популяции, то этот процесс идет медленнее. Мы вычислили величину коэффициента инбридинга для ляпинской и обеих сосьвипскнх групп манси. Для вычислений использовались формулы F, основанные на различиях в эмпирпческих (H f ) и ожидаемых (Я0) частотах гетерозиготных форм, которые возникают в популяциях при инбридинге. Hf=H0 + FH0. Вторая формула исходит из частот и гетеро- (Н) и гомозиготпых (DR)
4 DR — H*
форм F = /,/>/?_ //г л_ 2н • По-видимому, этот же результат, а именно превышение эмпирических частот гомозигот над теоретическими, должен быть и при действии генетико-автоматических процессов.
Для расчета использована система MN, где благодаря наличию двух сывороток выявляются гомо- и гетерозиготные формы.
Из табл. 1, где приводятся частоты групп крови, видно, что в Двух группах из трех, ляпинской и верхпесосьвпнской, ожидаемые частоты гомозигот больше, чем найдепо в действительности, в противоположность тому, что должно было бы быть при наличии инбридинга и при действии генетико-автоматических процессов. То же самое можно сказать и о системе Л ВО, где ожидаемая частота группы 0 выше, чем эмпирическая. Поэтому и величина F в этих группах имеет отрицательные величины ц равна — 0,0138 и —0,1242. В среднесосьвинской же популяции которую в известном смысле можно считать сборной, так как она включает уроженцев большого числа поселков, сильно удаленных друг от друга, F имеет положительную величину и равно 0,0206. Возможно, что различия в величинах F в исследованных группах носят случайный характер. Во всяком случае можно считать, что действие инбридинга и генетико-автоматических процессов, проявляющееся в возрастании частот гомозиготных форм, в нашем материале не улавливается. Может быть, влияние этих факторов затушевывается действием других явлений, таких, как отбор. Возможно, какую-то роль в том, что действие инбридинга не проявляется, играло фратрнальное деление изученных популяций в прошлом пли же тот факт, что Не все браки заключаются внутри популяций и идет пебольшой, по постоянный приток генов извне и отток, очевидно, тоже.
Различие исследованных групп мапси объясняется, по-видимому, прежде всего различием исходных популяций и их дальнейших исторических судеб. Придавать же большое значение действию гепетико-автоматическпх процессов и инбридинга в данном случае, видимо, нет оснований.
Таким образом, березовских манси можно разделить на три обособленные брачными кругами группы: ляппнскую, верхне-сосьвипскую и среднесосьвннскую. Все они по частотам гена q системы АВО приближаются к поволжским народам.
В ляпинской популяции с ее более низкими частотами генов q и т, по-видимому, прослеживается влияние палеоазиатских народов. Верхнесосьвппская группа обнаруживает примесь южно-сибирских монголоидных элементов с их более высокими частотами генов q, тп и более низким Pi и d. Средпесосьвинская популяция занимает промежуточное положение между двумя другими по большинству генов крови.
Несмотря па то что подавляющее число браков манси заключают внутри популяции и следовало бы ожидать нарастания частот гомозиготных форм, благодаря действию генетико-автоматических процессов и влиянию инбридинга, на наших материалах этого факта не наблюдается. Поэтому большого значения действию этих факторов в возникновении различий между популяциями в данном случае придавать не следует.
Литература
Лржелас Л. К. Выяплеппе агглютиногенов системы Льюис n кропи жителей Москвы. «Вопросы антропологии», 1965, вып. 19.
Бахрушин С. В. Остяцкие п вогульские княжества в XVI—XVII вв. Л., 1935.
ЩМаШлай А. Г. Системы ABO, Rh и Cell-Cellano по данным о первичных донорах г. Москвы. «Труды VII Международного конгресса антропологических и этнографических наук», т. 1. М., 1968.
Беневоленская Ю. Д. О характере полиморфизма популяции человека по системе резус. Сб. «Антропология и геногеография». М., 1974.
Бунак В. В. Антропологический тип черемис. «Русский антропологический журнал», т. 13, вып. 3–4. М., 1924.
Бунак В. В. Русское население n Забайкалье. «Антропологический сборник IV». «Труды Института этнографии АН СССР», новая серия, т. 82, 1963.
Бунак В. В. Гено-географнческие зоны Восточной Европы, выделяемые по факторам АВО. «Вопросы антропологии», 1969, № 32.
Гохман И. И. К антропологии елогуйских кетов. «Краткие сообщения Института этнографии АН СССР», вып. XXXVIII. М., 1959.
Дебец Г. Ф. Проблема заселения Северо-Западной Сибири, по данным палеоантропологии. «Краткие сообщения Института истории материальной культуры», вып. IX. М.—Л., 1941.
Золотарева И. М. Распределение групп крови у народов Северной Сибири. «Доклады па VII Международном конгресс антропологических и этнографических наук». М., 1964.
Золотарева //. М.. Башлай А. Г. Серологические исследования в Якутии. «Советская этнография», 1968, № 1.
Лашук Л. П. Этническая история Печорского края. «Краткие сообщения Института этнографии АН СССР», вып. XXI, 1956.
Левин М. Г. Группа крови у чукчей и эскимосов. «Советская этнография», 1958, № 5.
Левин М. Г. Новые данные по группам крови у эскимосов и ламутов. «Советская этнография», 1959, № 3.
Новицкий Г. Краткое описание о народе остяцком. СПб., 1884.
Патканов С. К. Статистические данные, показывающие племенной состав населения Сибири, язык и роды инородцев, т. I. СПб., 1912.
Патканов С. К. Список народностей Сибири. Пг., 1923.
Рычков Ю. Г. Некоторые аспекты серологических исследований в антропологии. «Вопросы антропологии», 1965, вып. 19.
Рычков Ю. Г. Особенности серологической дифференциации народов Сибири. «Вопросы антропологии», 1965, вып. 21.
Рычков Ю. Г., Перевозчиков II. В.. Шереметьева В. А., Волкова Т. В., Башлай А. Г. К популяционной генетике коренного населения Сибири. Восточные Саяны. «Вопросы антропологии», 1969, № 31.
Трофимова Т. А. и Чебоксаров H. Н. Антропологическое изучение манси. «Краткие сообщения Института истории материальной культуры», вып. IX. М—Л.. 1941.
Умнова М. А., Прокоп О., Пискунова Т. М., Самусева Г. С., Ича-ловска Т. А., Прозоровская Г. П. Распределение факторов крови у населения Москвы. Доклады на VII Международном конгрессе антропологических и этнографических наук». М., 1964.
Чернецов В. Н. «Фратриальное устройство обско-югорского общества», «Советская этнография», 1939, № II.
Чернецов В. Н. Очерк этногенеза обских югоров. «Краткие сообщения Института истории материальной культуры», вып. IX. М.— JL, 1941.
Штерн К. Основы генетики человека. М., 1965.
Li C. C. Population Genetics. N. Y., 1954.
Schwidetzkl I.. Walter H.. Knussmann R. Die neue Bussenknnde mit Beilragen von A. Bemane. Stuttgart, 1962.
Knussmann R. Moderne statistische Verfahren in der Hassenkunde. В кн.: /. Schwidetzky (Hrsg.). Die neue Rassenkunde. Stuttgart, 1962.
Более 800 000 книг и аудиокниг! 📚
Получи 2 месяца Литрес Подписки в подарок и наслаждайся неограниченным чтением
ПОЛУЧИТЬ ПОДАРОК