О генетической структуре популяций лопарей
Л. В. Эрикссон
Лопарей всегда было очень мало, вероятно, их число никогда не превышало сегодняшнее, т. е. приблизительно 35 тыс. на всем Севере. Около 30 тыс. из них живет в Финляндии. Предполагается, что 2 тыс. лет назад лопари занимали всю Северную Фенноскандию (почти всю Финляндию и Восточную Карелию). За последние 2 тыс. лет они проживают в глухих и гористых районах северных окраин Фенноскандии.
Еще 400–500 лет назад очень много лопарей было в восточных районах Финляндии. Утсойки и Инари — две самые северные общины Финляндии, где в настоящее время живет большинство финских лопарей.
За прошедшие пять лет было исследовано более 1200 лопарей из района Инари. Около 350 из них — сколтские лопари — сколты Севеттиярви, проживавшие ранее в деревне Суенйел, около 250 — лопари-сколты Неллима, которые раньше жили на р. Пасвик в районе Петсамо (Nickul, 1948; Lewin, Rundgren, Forsius, Eriksson, 1971). Вокруг озера Инари в настоящее время живет ровно 500 лопарей-рыболовов, которые долгое время не были изолированы. В западных районах общины Инари живет 450 горных лопарей.
Еще 100 лет назад в Инари было очень мало финнов, сейчас же финны составляют большинство, и лишь 20% всего населения, насчитывающего в настоящее время здесь более 7 тыс. человек,— лопари. Эти цифры свидетельствуют о том, что в общине Инари идет значительная ассимиляция финнами лопарей и разрушение их изолятов (Asp, 1966). В рамках Международной Биологической Программы по скандинавской секции адаптации человека в северной части Финляндии среди лопарей работало 130 исследователей из 16 стран, представлявших различные области науки. Были проведены как поперечные, так и продольные в плане организации выборки исследования, особенно среди лопарей-сколтов. Определялись изменения прироста, возрастная инволюция и динамика различных биохимических переменных величин в пубертатный период (Lewin and Eriksson, 1970; Lehmann, 1971).
Как правило, исследовались целые семьи. Для этого выбирались лица, которые по генеалогическим данным и языковым признакам были чистокровными лопарями. В Инари было обследовано более 95% оседлых лопарей-сколтов и более 50% лопарей-рыболовов и горных лопарей. Для вычисления частот генов были отобраны лица, у которых среди предков было не менее 75% чистокровных лопарей.
Мультифакторные признаки у лопарей
Что касается морфологических признаков, то лопари обнаруживают как «восточные», так и «западные» признаки. Цвет радужной оболочки у них намного темнее, чем у окружающих популяций, которые относятся к самым светлым народам в мире. Однако цвет глаз и волос не такой темный, как у азиатских народов. Так примерно у 30–50% лопарей глаза голубые (Forsius et ai., 1970) и даже у чистокровных детей лопарей (особенно у лопарей-скол-тов) часто встречаются светлая кожа и очень светлые волосы (рис. 1).
Другие признаки, отмеченные у лопарей, которые рассматриваются как «восточные», — слабый рост бороды, особенно на щеках, и слабый или даже совершенно отсутствующий волосяной покров на теле. В противоположность другим народам Северной Европы лопари-мужчины лысеют довольно редко, их волосы седеют или редко пли сравнительно поздно. Как и у других европей-
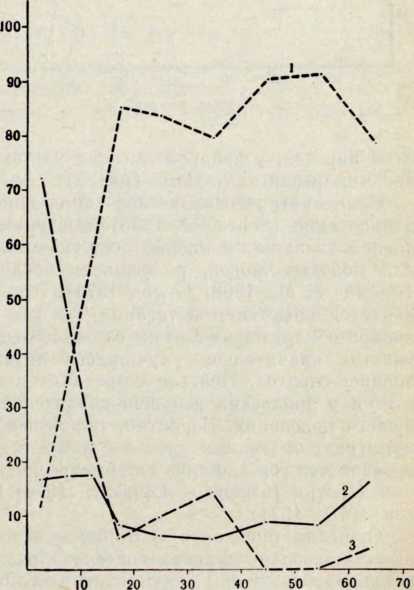
Возраст в годах
Рис. 1. Цвет волос лопарей-сколтов из Севеттиярви (но Шкале Фишера — Заллера). Обследовано 162 мужчины и 157 женщин
1 — U—Т (каштаново-черные волосы),
2 — Р—Т (каштановые полосы — шатены),
3 — А—О (светло-русые волосы — блондины)
Рис. 2. Обволошенность сред, ней фаланги у лопарей-сколтов Севсттиярвн и гренландских эскимосов
1 — лопари-сколты из Се-
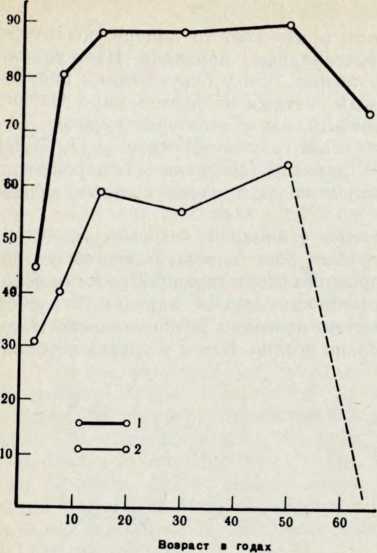
веттиярве, п=336,
2 — эскимосы северо-запад.
ной Гренландии, n= 1 44
ских народов, у лопарей высокая частота волосяного покрова на средних фалангах пальцев (рис. 2).
Соматометрические исследования показывают, что широкое и низкое лицо (Schreiner, 1935), выступающие скулы, высокий головной указатель и низкий рост значительно меньше характерны для молодых людей, родившихся после второй мировой войпы (Forsius et ai., 1968; Lewin, 1971). Это, по всей вероятности, не является следствием метисации, так как и чистокровные молодые лопари обнаруживают те же самые изменения. За последние десятилетия значительно улучшилось питание лопарей и особенно лопарей-сколтов. Половое созревание в настоящее время фиксируется у лопарских девушек значительно раньше, чем у предыдущего поколения. Вероятно, так называемый процесс акцелерации (ранняя половая зрелость и более высокий рост) у лопарей проявляется гораздо ярче или быстрее, чем у окружающего населения (Lewin, J?rgens et Louekari, 1970; Lewin, Nickul and Eriksson, 1970–1971).
Один из признаков, который рассматривается как «восточный»—довольно сильная конгенитальная аплазия некоторых коренных зубов. Вторые премоляры часто отсутствуют, это особенно характерно для лопарей-сколтов. Им присуща также несколько более высокая частота лопатообразных форм резцов. Бугорок Капабеллн, который у монголов отсутствует, встречается у лопарей-сколтов довольно часто. Что касается морфологии зубов, то одонтологи считают, что у лопарей-сколтов имеются и европейские и азиатские признаки (Zoubov, 1972).
Лопарям присущи также многие «западные» морфологические Признаки. Рефлектометрические исследования показывают, что кожа у лопарей пе такая желтая, как утверждали раньше. Монгольское пятно, характерное для детей азиатских народов (приблизительно в 80%), у лопарей встречается редко. Болосы у них не жесткие в противоположность большинству азиатских народов.
У финских лопарей специалисты по дерматоглифике выявляют как «восточные», так п «западные» черты (Lehmann et ai., 1970; Chit, 1972). Как показала К. Марк (1972), «восточные» черты («указатель монголоидности») встречаются у лопарей значительно чаще, чем у финнов и шведов Финляндии.
Моногенные признаки у лопарей
Частоты групп крови
По некоторым мономерным признакам популяции лопарей очень гетерогенны. Между отдельными популяциями имеются существенные различия, особенно в группах крови системы АВО (табл. 1).
Лопари-рыболовы отличаются от других популяций лопарей чособепно наглядно, для них характерна высокая частота гена 0 и почти полное отсутствие гена В. Общий признак для всех популяций лопарей — очень высокая частота гена А2. У лопарей-скол-тов частота генов В и М выше, чем у других лопарей.
Индивидуумы с отрицательным резус-фактором встречаются У лопарей редко. У финских лопарей частота гена d составляет примерно четвертую часть частоты этого гена у окружающих популяций. Ген имеет у лопарей высокую частоту, п эта частота возрастает в восточном направлении.
Частота аллеля Duffy (Fya) высокая, особенно у западных лопарей. Аллель Kidd (Jka) имеет низкую частоту у всех до сих Пор исследованных лопарей. Антиген Diego (Dia), характерный только для монголоидной расы, полностью отсутствует у скандинавских лопарей (число обследованных более 1000 человек) и финнов, но нередко встречается у марийского населения по излучине Волги.
Частота лейкоцитного антигена (HL—.4) и его комбинации У 300 исследованных лопарей-сколтов отличаются от частот в окружающих популяциях (Tiilikainen, Eriksson, Macqueen et Amos, 1972).
Ген Шведы и норвежцы финны Шведские лопари * ®5I«:г с — о X Ч Горные лопари Инари Лопари-рыболовыИнари Лопарн-сколты 3? А 2-<4| 0,4 0,4 6–7 1–2 3 3 0,5–2 0,2 Аг 7 10 32–37 25–36 31 20 14–16 4 В 7 13 2–3 9–14 7 0,4 14–33 18 MS 24 25 25 19–27 25 33 29–31 21 Ms 32 40 14 26–33 20 18 32–38 36 NS 8 8 20 15–18 10 И 5 6 Ns 35 28 42 30–32 45 38 26–35 38 r (cde) 38 34 20–29 16–19 10 6 9–10 38 ?H ЛГ 42 43 53–61 51–59 63 64 67–79 45 cw 2 2 4–10 4 2 5 0–1 2 p 51 45 37–39 31–53 38 43 26–36 45 Duffy /•//" 42 47 55 54–82 57 55 70–74 60 Keli K 4 2 0,3 6 6 0,2 1–2 2 Kidd Jk" 51 47 53–66 30 19 20–30 49 Diego Dia 0 0 6 0 0 0 0 1,3 * Данные по шведским и норвежским лопарям включают в себя результаты следующих исследований: Элипен и др., 1952; Элизен и др , 195П; Бекман и др., 1959; К<>рн-штад, 1972; Монн, 1969.Полиморфизм белка плазмы
Частоты генов белка плазмы также обнаруживают у лопарей гетерогенную картину. Для гена Gc2 характерны значительные колебания частот (рис. 3): у лопарей-рыболовов он имеет низкую частоту (3%), а у ближайших соседей лопарей-сколтов Неллима — высокую.
Хорошее совпадение наблюдаемых и ожидаемых частот готерозигот довольно неожиданно, если учесть, что эти популяции были более или менее изолированны и что в число обследованных входит много родственников, в том число самых близких. В системе //р наблюдается значительная внутренняя гетерогенность, но есть и относительные небольшие различия между отдельными популяциями лопарей (Eriksson et ai., 1970).
Медленная подтиповая компонента гена Нрх имеет высокую частоту у лопарей-сколтов. В настоящее время они являются единственной популяцией лопарей, у которой исследовалась эта подтпповая компонента (Ehnholm et Eriksson, 1969). Трансферинный ген Dchi имеет у финнов, по сравнению с другими европейцами, высокую частоту, а у большинства исследованных лопарей он встречается редко.
Ген С'З2 имеет высокую частоту в третьем компоненте у фпЯ-
о а • б
--1–1-1–1-1–1-1–1-г
0.1
01 23456789 10
Рис. 3. Пропорции гетеро:шгот в системах G с и Нр
а — Gc2, б — Нрг, 1 — сколты Севеттиярви (п=327), 2 — сколты Неллима (п= 131), 3— лопари-рыболовы (п=207), 4 — горные лопари (п=125), 5 — потомки лопареи-рыболовов и горных лопарей (п=77), 6 — лопари-сколты, (п=26), 7 — сколтские финны (п=97), 8 — лапландские финны (п=88)
ских лопарей (Berg et ai., 1972). Из наследственных вариант антитрипсина альфа в сыворотке более низки частоты других типов Р(, чем MM (Fagerhol et ai., 1969).
Аллотипы гамма-глобулина у финских лопарей
Gm (1) имеет у европейцев частоту приблизительно 50–00%, у негров, монголоидных народов и австралийцев-аборигепов — 100%. У лопарей-рыболовов в Инари самая высокая из до сих пор зафиксированных в Европе частот — около 80%. Лопари-сколты имеют низкие частоты Gm (1), которые выражаются значениями между 44% (Неллим) и 59% (Севеттиярви).
Gm (2) имеет у лопарей-сколтов низкую частоту (Севетти-ярви 3%, Неллим 7%), для финнов же характерна самая высокая из зафиксированных частот Gm (2). Другие финно-скандинавские популяции имеют также высокую частоту фенотипа Gm (2). Следует отметить, что в популяции марийцев частота Gm (2) составляет всего 11%, что примерно втрое ниже частоты Gm (2) у финнов. У венгров еще более низкие частоты Gm (2) (5–8%). Лопари-сколты имеют самую низкую из зафиксированных частот Gm (2) в Европе (3,2%). По частотам Gm (5) финские лопари также обнаруживают очень гетерогенную картину. Лопари-рыболовы имеют самую низкую из зафиксированных частот Gm(5) в Европе (70%), у лопарей-сколтов наблюдается очень высокая частота Gm(5) (93–95%).
Все исследованные популяции полиморфны по антигену Inv Inv1. У большинства европеоидных народов частота гена Inv значительно ниже, чем у индейцев в Южной Америке, меланезийцев и т. д. Восточноазиатские народы имеют частоту Inv1 около 30%, Финно-угорские народы, такие, как марийцы, венгры и финны, имеют частоты Inv1 ниже 5%. У финских лопарей частоты Inv1 в 3–5 раз выше. Вероятно, в отношении Inv1 они очень резко отличаются от окружающих народов.
Полиморфизм ферментов
У финских лопарей, а особенно у лопарей-рыболовов и горных лопарей, частота генов RGM и ADA2 является самой высокой из до сих пор известных частот этих генов. В других популяциях финно-угорской языковой группы, как, например, у финнов и марийцев, по сравнению с другими европейцами частоты гена ADA2 тоже высокие. Все до сих пор исследованные лопари Норвегии, Швеции и Финляндии имеют низкие частоты АК2. Аллель АК2 полностью отсутствует у лопарей-сколтов.
У лопарей отмечается очень высокая частота гена кислой фосфатазы в эритроцитах; высокая частота этого гена характерна также и для финнов. Эти частоты гена Р° самые высокие из до сих пор известных во всем мире (Eriksson, 1968).
Вариант супероксидазы (Superoxidismutasvariante), называемый также вариантой индофенолоксидазы, тетразольной оксидазы или вариантной «white patch», можно увидеть в окрашенном тетразолием крахмальном геле в виде обесцвеченных участков. Этот наследственный вариант имеет высокую частоту у лопарей-сколтов Неллнма. Его появление, видимо, вызвано потоком финских генов, так как он полностью отсутствует у всех остальных исследованных популяций лопарей. Складывается впечатление, что этот «частный вариант энзима» имеет исключительно низкую частоту у всех до сих пор исследованных популяций, кроме финнов. У финнов же он является, по всей вероятности, полиморфным признаком.
Компонент С5 псевдохолннэстеразы имеет высокую частоту у лопарей-сколтов (12–15%), у финнов его частота низкая (всего 3–5%). Сфера распространения дибукапнового резистентного варианта псевдохолннэстеразы хорошо согласуется с данными, полученными в других популяциях (E“ =0,0070–0,02338) (Singh et ai., 1971).
Другие наследственные признаки
Частоты индивидуумов, не имеющих вкусовой чувствительности к фенилтиокарбамиду (РТС), составляет у лопарей приблизительно 10%, у финнов эта частота в 2–3 раза выше. Исключением являются лопари-сколты: приблизительно 29% из них лишены вкусовой чувствительности к PTC (Eriksson, Fellman, For sius et Lehmann, 1970).
Создается впечатление, что аномалии цветового зрения (про-ханопия, дейтераноиия) имеют у лопарей несколько более низкую частоту, чем у финнов (5 или 8% у мужчин) (Forsius, jjriksson et Fellmann, 1968; Forsius, Luukka et Eriksson, 1971).
По всей вероятности, у лопарей высока частота (примерно 65%) быстрых инактиваторов изониацпда (INH). Частота их у других европейцев составляет 34–45%, У азиатских народов и эскимосов она достигает 70–90% (Tiitinen et ai., 1968). Утверяедают, что быстрая пнактпвацня 1NH в Азии обнаруживает к северу более высокую частоту. Но вопрос, не связано ли это с фактором среды, пока остается открытым.
Таблица 2. Типы ушной серы у лопарей Популяции n сухапсера,°/о W Лопари-сколты Севеттнярви .... 254 11,41 0,059 Лопари-сколты Неллима...... 111 12,61 0,065 Лопари-рыболовы ........ 146 5,48 0,028 Горные лопари ..........Потомки лопарей-рыболовов и гор 101 2,97 0,015 ных лопарей ........... 63 6,35 0,032 Потомки лопарей-сколтов п финнов 78 7,69 0,039 Потомки лопарей и финнов .... 73 4,11 0,021Частота сухого типа ушной серы (ww) очень высока у азиатских народов (свыше 80–90%). У европейцев этот фенотип встречается относительно редко (всего около 5–10%), чему соответствуют данные, полученные в результате исследования лопарских популяций (табл. 2).
Патологические признаки у лопарей
Хотя пока нельзя назвать точные цифры, тем не менее складывается впечатление, что для финских лопарей мало характерны такие заболевания, как холецистит, аппендицит, ожирение, варикозные расширения вен и гипертония. У лопарей наблюдается высокая частота случаев плоскостопия u врожденного вывиха тазобедренного сустава. В некоторых популяциях частота этой Патологии примерно в 10–30 раз выше, чем у окружающих позиций (табл. 3).
Очень трудно объяснить, чем вызвана у лопарей высокая частота плоскостопия и вывиха тазобедренного сустава, в то время как хождение пешком — основной способ их передвижения. Это кажется чем-то вроде парадоксального отбора или случайного зйфа генов.
В прежние времена матери очень туго пеленали грудных детей и пользовались специальными лопарскими колыбелями, на-
зываемыми гитка или комсио. В настоящее время медицинские сестры и врачи запретили ими пользоваться, так как грудные дети не могли в них двигаться. Частота врожденного вывиха тазобедренного сустава в последних поколениях стала значительно ниже, по крайней мере у лопарей, проживающих в районе Инари.
Типичные для лопарей признаки
При исследовании небольших и изолированных групп населения, какими являются лопари, не следует делать далеко идущих обобщений. Эффект родоначальника и случайный дрейф генов имели, по-видимому, в этих случаях большое значение. Лопари-сколты были немногочисленной изолированной окраинной популяцией (рис. 4), которая по многим признакам отличается от лопарских популяций в основной зоне обитания лопарей.
Однако существуют некоторые генетические характерные признаки, которые присущи только лопарям и которые отличают их от окружающих популяций.
Лопари отличаются от всех до сих нор исследованных соседних популяций исключительно высокой частотой гена А2. Общими почти для всех исследованных популяций лопарей являются высокие частоты генов Аг, RGM2, ADA2 и Inv1 и низкие частоты генов ff, М, Pi, Gc2 и АК2.
Лопари обнаруживают сходство с восточными популяциями, для тех и для других характерна низкая частота хромосомы г. .ffi-хромосомы и аллель Duffy (Fya) имеют высокие частоты. У всех до сих пор исследованных лопарей отмечается низкая частота аллелей К, Kidd (Jka) и несекреторного аллеля se.
Имеются доказательства присутствия у лопарей-сколтов «восточного» аллеля гамма-глобулина Gm1'13- 15, но некоторые другие «восточные» признаки Gm отсутствуют, а аллели Gm3'5- 13, 14>
400-j
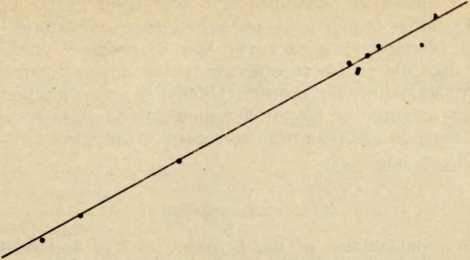
10 -I-1------1-----—»—----1-
1500 1600 1700 1800 1900
Рис. 4. Средний ежегодный прирост численности лопарей-сколтов Суениела
(Севеттнярвн)
На оси ординат — размер популяции на логарифмической шкале, на оси абсцисс — годы. Средний ежегодный прирост равен примерно 0,63%
200-
100-
60-
40-
20-
характерные для европеоидных популяций, имеют высокую частоту. Для финских лопарей характерна значительно более высокая частота аллеля Inv1 (25–30%) по сравнению с другими североевропейскими популяциями (5–10%). Частота гена Inv1 у восточноазиатских популяций высокая (30–40%).
Поток генов от финнов к лопарям
V финских лопарей гены В, cde (r) и АК2 имеют очень низкие частоты. Если предположить, что в исходных популяциях лопарей эти гены отсутствовали и что наличие их у нынешних лопарей объясняется потоком генов от окружающих популяций, то величину этого потока можно оценить приблизительно в 20–50% в зависимости от того, какие гены исследуются.
Популяционно-генетические исследования подтверждают, что популяции лопарей во многих отношениях исключительно своеобразны. Ярко выраженная теистическая гетерогенность отдельных популяций указывает на то, что большинство экстремальных частот генов можно объяснить «эффектом родоначальника» и «случайным генетическим дрейфом» в небольших и изолированных популяциях, которые жили в суровых условиях и часто на грани вымирания. Такие стихийные бедствия, как эпидемии и голод, могли вызвать огромные изменения в структуре популяций, 4 результате чего последствия дифференциальной плодовитости и дифференциальной смертности были бы особенно наглядны.
Популяции лопарей очень долго были изолированы и разбросаны на огромной территории. Тем не менее существуют наследственные признаки, которые все еще имеют (по крайней мере у популяций лопарей в их основной зоне обитания) четко выраженную «лопарскую» частоту. Это тем более удивительно, что должен был иметь место значительный поток генов от большинства окружающих популяций. Очевидно, по крайней мере некоторые из предковых групп лопарей имели какие-то экстремальные частоты генов, влияние которых у современных лопарей заметно до сих пор.
Литература
Asp Е. The finnicization of the Lapps. A case of acculturation. «Annales Univ. Turkuensis», Ser. B, t. 100. Turku, 1966.
Berg K., Arvilommt H. and Eriksson A. W. C3 polymorphism among Finnish Lapps. «Human Herediy», 1972 (9 in proof).
Chit H. L. ?ber das Hautleistensystem der Bev?lkerung Finnlands. «Annales Academiae Scientiarum Fennicae», AV, N 151, 1972.
Ehnholm C. and Eriksson A. W. Haptoglobin subtypes among Finnish Skolt Lapps. «Ann. Med. exp. Fenn.», 1969, N 47.
Eriksson A. W.. Fellman Forsius II. and Lehmann W. Phenylthiocarbamide tasting ability among Lapps and Finns. «Human Heredity», 1970, N 20. Eriksson A. W., Kirjarinta M., Fellman Ehnholm Ch., Lehmann W. ocli Forsius H. Kvalitativa och kvantitativa haptoglobinunders?kningar bland samer. «Nord. Med.», 1970, N 84.
Fagerhol M. K.. Eriksson A. W. and Monn E. Serum Pi types in some Lappish and Finnish populations. «Uuman Heredity», 1969, N 19. Forsius H., Eriksson A. W. and Fellman J. Colour blindness in Finland. 1. Frequency of protans and deutans. 2. Family studies on minor deviations of the normal mit matching point using the anomaloscope. «Acta ophthalmologica», Kobenhavn, 1968, N 46.
Forsius H., Lehmann W., Fellman J. and Eriksson A. W. Iris pigmentation of Skolt Lapps and Finns. «The Scandinavian Journal of Clinical and Laboratory Investigation», 1968, N 21, Suppl. 101.
Forsius H.. Luukka //. and Eriksson A. W. Ophthalmogenetic studies on the Skolt Lapps. «Acta ophthal.». Kobenhavn, 1971, 49.
Forsius H.. Luuka H.. Lehmann W., Fellman J. och Eriksson A. W. Irisf?gr, korneabrytningsf?rm?ga och korneatjocklek bland skoltsamer och finnar. «Nord. Med.», 1970, N 84.
Getz B. The hip joint in Lapps and its bearing on the prob'um of congenital dislocation. «Acta orthopadica scand.», 1955, Suppl. 18.
Kornstad L. Distribution of the blood groups of the Norwegian Lapps. «American Journal of physical Anthropology», 1972, N 36.
Laurent L. E. Congenital dislocation of the hip. «Acta chir. scand.», 1953, Suppl. 179.
Lehmann W. (Koordinator). Der Mensch in der Arktis. «Anthropologischer Anzeiger», 1971, N 33.
Lehmann W.. Eriksson A. И^., J?rgens H. W. och Forsius H. Iludlinjesyste-met hos skoltsamer. «Nord. Med.», 1970, N 83.
Lewin T. Introduction to the biological characteristics of the Skolt Lapps. «Proc. Finn. Dent. Soc.», 1971, N 67, Suppl. 1.
Lewin T. and Eriksson A. W. The Scandinavian International Biological Program, section for Human Adaptability, IBP/HA. «Arctic Anthropology», 1970, N 7.
Lewin Т.. Jurgens H. W. and Louekari L. Secular trend in stature of adult Skolt Lapps between 1915, 1934 and 1968. Studies of stature change4 on population and family levels of a genetic isolate. «Arctic Anthropology», 1970, N 7.
Lewin Т., Mickul K. and Eriksson A. W. Stature and ageing of the Skolt Lapps. «Acta morph, neer-scand.», 1970–1971, N 8.
leivin Т., Rundgren A., Forsius H. and Eriksson A. W. Demography of the Skolt Lapps in Northern Finland. «Proc. Finn. Dent. Soc.», 1971, N 67, Suppi. 1.
Mark K. Anthropologische Eigenschaften der Bev?lkerung Finnlands. «Annales Academiae Scientiarum Fennicae», AV, 152, 1972.
Mellbin T. The children of Swedish nomad Lapps. A study of their health, growth and development. «Acta paediat.» (Uppsala), 1962, N 51, Suppl. 131.
N??t?nen E. K. ?ber die Anthropologie der Lappen in Suomi. «Annales Academicae Scientiarum Fennicae». Ser. A, t. 47, N 2, 1936.
Sickul K. The Skolt Lapp community Suenjelsijd. During the year 1938. «Almqvist & Wiksell». Uppsala, 1948.
Palmen К. Preluxation of the hip joint. Diagnosis and treatment in the newborn and the diagnosis of congenial dislocation of the hip joint in Sweden during the years 1948–1960. «Acta paediat.» (Uppsala), 1961, N 50, Suppl. 129.
Sehreiner K. E. Zur Osteologie der Lappen. Ser. В, XVIII, 1 und 2. Oslo, 1935.
Singh S., Jensen M., Goedde H. W., Lehmann W., Py?r?l? K. and Eriksson A. W. Pseudocholinesterase polymorphism among Lapp populations in Finland. «Human Genetics», 1971. N 12.
Subow A. A. Einige Angaben der Dentalen anthropologie ?ber die Bev?lkerung Finnlands. «Annales Academiae Scientiarum Fennicae», AV, N 150, 1972.
Tiilikainen A., Eriksson A. W., MacQueen 1. M. and Amos D. B. The HL-A system in the Skolt Lapp population. «Proc. Fifth Int. Histocompatibility Workshop Conf.». Evian, 1972.
Tiitinen H., Mattiia M. J. and Eriksson A. W. Comparison of the isoniazid inactivation in Finns and Lapps. «Ann. Med. intern. Fenn.», 1968, N 57.
Wessel A. В. Laaghalte slegter i Finmarken. «T. norske Laegeforen», 1918, N 38.
Более 800 000 книг и аудиокниг! 📚
Получи 2 месяца Литрес Подписки в подарок и наслаждайся неограниченным чтением
ПОЛУЧИТЬ ПОДАРОК