Распределение различных генетических маркеров в Финляндии и проявление их в Эстонии и Венгрии
X. P. Неванлинна
Прежде чем представить собственные данные по финнам, эстонцам и венграм, мне хотелось бы более детально обсудить некоторые факторы, которые могли бы влиять или действительно влияют на вариабельность в человеческих популяциях, что является темой нашего симпозиума.
Необходимо также обратить внимание на некоторые качества методов, использовавшихся для измерения биологических характеристик человека. Внимание будет сконцентрировано на оценке методов, примененных для определения полиморфизма человека, и только коротко будет упомянуто о факторах, связанных с ошибками в измерении физических характеристик, т. е. физической антропологии.
Полиморфизм человека. Согласно Форду, ген называют полиморфным, если частота редкой аллели выше ожидаемой при учете роли только одних мутаций. На практике к полиморфным относятся гены с частотой редкого аллеля выше 1 %. Существует множество генов, которые встречаются в одних популяциях с частотой намного большей, чем приведенная выше, но совершенно отсутствуют в других. Уже само определение полиморфизма исключает мутацию как причину распространения этих генов, и для обсуждения остаются два известных фактора эволюции: случайный генетический дрейф и отбор.
Необходимо подчеркнуть одно обстоятельство. Если для характеристики популяций человека применяется понятие полиморфизм, то нужно учитывать, что гены, а не признаки передаются из поколения в поколение. Хотя это и должно быть более чем очевидно для любого, обладающего достаточными знаниями основ генетики, тем не менее даже в последние годы нередко можно видеть карты, таблицы, графики, где для иллюстрации и даже для сравнений между различными популяциями вместо частоты гена учитывают изменения фенотипа. Если поступать таким образом, то теряется важное преимущество — возможность правильной оценки.
Для пояснения возьмем два примера. Китайцы иногда рассматриваются как популяция, у которой «отсутствует Rh — отрицательный ген». К этому неправильному толкованию могло привести то обстоятельство, что, по-видимому, все китайцы Rh — положительны. На самом же деле, частота гена rh (Rh — отрицательного гена) около 0,05 (5%), например, среди китайцев, проживающих в США. Этой частоте соответствует один Rh — отрицательный индивидуум из 400. Соответственно фенотипическая частота в 20 раз меньше частоты гена.
Распределение фенотипа и генотипа редкого характера (/?/(-отрицательный)
Частота гена г .... Частота (приблизительно) 0,05(5%) 0,10 (10%)
Частота Л/(-отрнцателыюго=г2 . . . 0,0025 (1 : 400)
Данный пример относится к рецессивному гену, где разница между частотой гена и проявлением фенотипа огромная. Тем не менее это положение можно будет применить и к доминантным генам, если они будут весьма распространенными. Напомню, что частота группы крови Р* среди популяции составляет 75%. Если не знать (или если не помнить) о типе ее наследования, о том, что доминирует геп р (геп Р — ), тогда тот факт, что частота гена Р составляет только 0,50 (50%), может привести к совершенно неправильной оценке этого рода информации.
Распределение фенотипа и генотипа общего характера (Р)
Частота гена Р .........
0,50
0,75
Частота Р положительного—Р2-)-2Р7,
Случайный генетический дрейф. В любой ограниченной популяции — а всякая человеческая популяция имеет ограниченные размеры — частота всех аллеломорфных генов будет случайно колебаться от поколения к поколению. Эти колебания зависят от трех факторов: эффективного размера популяции (эффективный размер составляет приблизительно одну треть от общего), частоты гена n числа поколений. Конечным результатом явится утрата либо фиксация всех генов. В настоящее время в человеческих расах или в целых нациях из-за их огромного размера автохтонные процессы оказывают небольшое влияние на частоту встречаемости генов, но дрейф мог оказывать существенное влияние на ранних стадиях, в ранние исторические эпохи па те гены, которые наблюдаются теперь.
Хотя генетический дрейф не изменяет генные частоты в больших популяциях, он безусловно имеет значение для малых популяций. Это относится не только к примитивным популяциям с малым эффективным размером, но и к сельским популяциям развивающихся стран. Локальное изменение частоты генов может повлиять на общие результаты при неправильно полученной выборке.
Для демонстрации действия дрейфов генов можно привести примеры одного из последних исследований сельской популяции Финляндии. На рисунках 1 и 2 представлены распределения Двух маркеров — группы крови В и варианта Dem трансферрина — в области центральной Финляндии, насчитывающей около 10 000 человек. Область подразделена на группы деревень, в каждой из которых проживает около 500 жителей. Значительные различия между отдельными деревнями не могли слишком влиять па результаты, скажем, популяционного исследования, как раз именно при неоднородной выборке. Табл. 1 дает различия между
Таблица 1. Генные частоты аллелей, принадлежащих к 10 различным локусам финноязычной популяции: по стране в целом, среднее значение и ряд соответствующих оценок, полученных в семи общинах Ген По стране в целом n — 5536 Крайние значения по общинам Средняя А 0,2134 0,1357–0,2530 0,1936 а2 0,0956 0,0706–0,1035 0,0381 в 0,1320 0,0932–0,2304 0,1550 0 0,5589 0,48–43—0,6175 0,5633 MS 0,2469 0,2035–0,2832 0,2427 MS 0,3956 0,3234–0,4356 0,3837 NS 0,0773 0,0516–0,1152 0,0790 NS 0,2801 0,1923–0,3646 0,2945 S 0,3241 0,2788–0,3817 0,3222 CDe 0,4082 0,3117–0,4413 0,3945 CD E 0,1837 0,1355–0,2492 0,2036 С “De 0,0203 0,0050–0,0366 0,0192 cDe 0,0373 0,0055–0,0714 0,0394 Cde 0,0114 0,0000–0,0455 0,0154 cdE 0,0011 0,0000–0,0047 0,0005 cde 0,3380 0,2797–0,3704 0,3273 K 0,0203 0,0084–0,0517 0,0271 P 0,4511 0,3639–0,4810 0,4287 Fya 0,4710 0,3669–0,6167 0,4945 Hpi 0,3810 0,2930–0,6232 0,4188 Gci 0,7947 0,7510–0,8294 0,7944 TIC 0,9779 0,9416–0,9919 0,9710 H(y- 1 0,0116 0,0020–0,0311 0,0117 Bi 0,0001 0,0000–0,0000 — B2 0,0009 0,0000–0,0058 0,0012 Dchi 0,0090 0,0000–0,0389 0,0156 Dfin 0,0005 0,0000–0,0031 0,0005 Nem Ts 0,4596 * 0,4069–0,5620 0,4797 * Данные получены Аллисоном и Неванлинной (1951 г.)большими популяционными группами внутри одной и той же нации. Частоты генов в семи сельских общинах Финляндии, насчитывающих до 10 000 жителей каждая, отличаются значительно больше, чем у финнов по сравнению с любой другой нацией. Поэтому в популяционных исследованиях все усилия должны быть направлены на получение соответствующей выборки.
То, что значительные локальные отклонения в Финляндия действительно зависят от случайного дрейфа, видно при сложении локальных выборок. Эти частоты генов очень близки суммарным финским частотам. В этой связи необходимо напомнить,
Рис. 1. Частоты группы крови В системы А ВО на территории центральной части Финляндии
1 — низкие частоты, 2 — средние частоты, 3 — высокие частоты
что до последнего времени было принято считать, что нарастание частот генов В и М с запада и юга на восток и север отражает различия в происхождении разных частей населения Финляндии. На самом же деле, характер распределения, наблюдавшийся более ранними аптропологами, по-видимому, отражает случайные отклонения, появляющиеся в несбалансированных выборках в популяции, гомогенной в другом отношении.
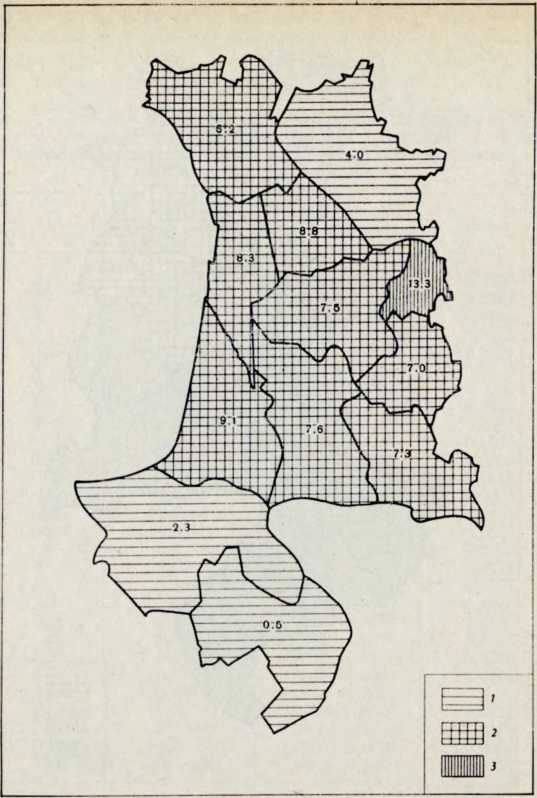
Рис. 2. Частоты трансферрина Drki на территории центральной части Финляндии 1 — низкие частоты, 2 — средние частоты, з — высокие частоты
Отбор. По-видимому, генетики все больше и больше принимают точку зрения, что генетический полиморфизм обеспечивает любой вид — включая человеческий — богатым источником для быстрого отбора, необходимого для любой эволюции, гораздо более быстрой, чем это может дать мутация. Уже это указывает на то, что так называемые нормальные маркеры, детерминированные полиморфными генами, не могут и не должны быть свободны от селекции. Можно только сожалеть, что практически ничего но известно о направлении, силе и механизме отбора и отношении групп крови и ферментного полиморфизма. Вообще вопрос о том, возникла дифференциация по этим маркерам под действием отбора или была обусловлена причинной взаимосвязью между этими маркерами и некоторыми болезнями, как предполагает в последней монографии профессор Н. П. Эфроимсоп, остается пока открытым.
По-видимому, пока не существует однозначного и простого пути для разрешения этой проблемы. Возможно, было большое число селективных факторов, которые дали современную картину полиморфизма. Можно предполагать, что силы отбора действовали медленно и различия между этническими группами, наблюдаемые в настоящее время, действительно вызваны различиями в происхождении этих групп пли популяций. Однако даже незначительное селективное преимущество могло решительным образом изменить распространение маркера. С другой стороны, быстрое увеличение числа известных маркерных генов позволяет надеяться, что часть из них могла появиться под действием сил отбора в такие далекие времена, что изучение пространственного распределения этих маркеров даст надежные данные антропологам.
Физическая антропология. Критические замечания в отношении использования факторов крови остаются в силе и тогда, когда речь идет о признаках со сложной генетической структурой. Вместе с тем необходимо подчеркнуть большое значение любых попыток представить общую картину распространения многих признаков человека. Использование же одного признака не дает возможности сделать сколько-нибудь существенные выводы.
Отмечу одну особенность действия отбора, когда речь идет о признаках, используемых в физической антропологии. Она заключается в сознательном выборе сочетающегося браком партнера. Результаты такого рода селекции ясно видны, например, в Финляндии или в любой популяции с ограниченной миграцией на относительно быстром образовании многочисленных субпопуляций, сильно отличающихся друг от друга морфологически.
Финны. Образование западной ветви угро-финской лингвистической популяционной группы — финнов — определено по времени и локализовано археологическими находками и лингвистическими данными, но исторических сведений пока еще недостаточно. Финны являются типичной пограничной популяцией со всеми ее особенностями: обширной площадью распространения, низкой плотностью населения, а также национальной и локальной изоляцией. Эффективная изоляция частично связана также с религиозными и лингвистическими барьерами. Финский изолят сформировался в древнее время (до 1200 г. до н. э.). Существуют лишь косвенные доказательства в пользу предположения, что финны сохранили лучше, чем более ранние ветви, свой генетический состав. Прежде чем перейти к этой важной детали, хотелось бы указать на некоторые типичные отличительные особенности финнов.
Влияние иммиграции на генофонд. В последнее десятилетие в нашей стране обнаружены разнообразные группы крови, сывороточные группы и наследственные болезни, которые вызываются либо неизвестными генами, либо такими, которые спорадически появляются в других частях света. Если не вдаваться особенно в детали, распространение многих подобных генов дает только одно возможное объяснение: они возникли в результате обогащения популяции, что вызвано ограниченной и медленной первоначальной иммиграцией. В табл. 2 схематично представлен
Таблица 2. Воздейстние раннем иммиграции на генетический дрейф(число иммигрантов в каждом поколении = 1.Популяции увеличивается приблизительно на 2% в год, т. е. удваивается через каждое поколение; <7 = 2)Поколение: I II III IV V VI ..........t 1 2 4 8 16 32 ..........2<-' 1 2 4 8 16 2<-2 1 2 4 8 2<-з I 2 4 ..........2 1 2 1 1 1 3 7 15 31 634
2

механизм подобного рода обогащения. На схеме продемонстрировано, что иммиграция происходила путем медленного притока некоторого числа иммигрантов, имеющихся в каждом поколении. Далее предполагается, что число иммигрантов увеличивается на 2% в год, это удваивает популяцию в каждом поколении. После какого-то числа поколений большая часть генофонда состоит из генов, происходящих от самых первых поселенцев. Естественно, что эффект будет лучше виден на чрезвычайно редких генах, которые ранние поселенцы приобрели из тысячи разновидностей генов. Часть накопленных генов могла быть получена па последних ступенях иммиграции. Вероятность этого явления возрастает с частотой индивидуальных генов. Этот механизм рассматривается здесь потому, что, по-видимому, он мог дать нашей популяции в прошлом гораздо больше типических черт, чем опа приобрела в течение последних двух тысячелетий. В заключение необходимо напомнить, что накопление всегда компенсируется потерями: финны в процессе этой многоступенчатой иммиграции также могли потерять значительную часть более редких генов.
Единообразие. Как уже указывалось, распределение распространенного генетического маркера в Финляндии удивительно однообразно, если оп изучается на уровне округа. Неравномерное распределение необычных генов, так же как и их ограниченное число, делает бесполезными поиски дальнейших различий. Единообразие может быть изучено и другим способом, т. е. рассмотрением распространенности редких генов. Это распространение имеет два характерных признака. Если частота маркера очень низкая, большинство генов локализовано в области первого поселения. Это указывает на то, что настоящая популяция имеет общее происхождение. Это предположение еще больше подкрепляется распространением четырех редких заболеваний, три из которых — типично финские. Расположение редких генов, по мнению автора, доказывает, что ранние западные поселенцы образовали также популяцию и на востоке. Являются ли сегодняшние карелы потомками древней популяции, жившей вокруг Ладоги, нельзя решить, используя генетические доказательства. Историки говорят, что первоначальная южнокарельская популяция, вероятно, двигалась на восток и что эта часть страны была вновь заселена с Запада и Севера. Этот факт очень хорошо сопоставляется с нашими данными.
Черты полиморфизма финнов. Кроме редких маркерных генов, имеется множество более распространенных генов, частота которых отличается от частоты, наблюдающейся в других странах. Это касается изученных до сих пор европейских популяций,
Таблица 3. Типичное распределение некоторых маркеров в Финляндии Ген Частота 1 Высокая частота м 0,64 Аг 0,10 Fya 0,47 8*1 0,80 Низкая частота rh 0,34 Kell 0,02для многих из них она является чрезвычайной. У финнов отмечается высокая частота ^2(^2), M, Fya fDuffy) и G'c (группспецифический компонент) (табл. 3). Существует несколько маркерных генов, которые также могут быть названы полиморфными,но их нет, например, в Скандинавских странах. К их числу относятся гены группы крови Ula и варианты трансферрипа ?o-i ц Don- Их частота достаточно высока, чтобы
Генетические маркеры, типичные для финнов
Частота
маркера
Группа крови Ul°............ 2,2°/с
А ...........1 : 6000
предположение, что они накоплены у ранних поселенцев, было бы невероятным. С другой стороны, у финнов обнаруживается чрезвычайно низкая частота гена Kell и rh(cde). Соответственно этому частота резус отрицательных лиц ниже, чем это до сих пор зарегистрировано для любой белой популяции, т. е. менее 12%. Наконец, необходимо знать, что частота группы крови В достаточно высока (q=0,1320), но ниже той, которая наблюдается у русских Ленинграда и Москвы.
Не вдаваясь в детали, необходимо обратить внимание на два обстоятельства. Во-первых, генный комплекс, свойственный финнам, характеризуется некоторыми не европейскими особенностями. Если исследуется его происхождение, необходимо искать популяции с частотой генов, сравнимой с этой характеристикой. Чтобы облегчить задачу, следует искать в прошлом смешение с популяциями, у которых имеется высокая частота M, Ai, UT, трансферрипа В0- и Dcm и отсутствует Kell и отрицательный Bh. Смешение такого рода генетического материала должно было быть достаточно сильным. Например, частота rh не может быть снижена с 40 до 34 при менее чем 20%-ном смешении с популяцией, у которой частота гена rh такая же, как у китайцев. Само собой разумеется, что смешение такого рода было, всего вероятнее, вызвано медленной, возможно, очень медленной миграцией гена из других популяций, обладающих подобным генетическим материалом.
Во-вторых, хотелось бы привлечь внимание к тому, что подобная генетическая модель позволяет сравнить финнов с эстонцами, чтобы выявить, какая из популяции лучше сохранила первоначальную модель балтийских финнов, или, иными словами, какая из двух популяций ближе к модели, наблюдаемой в других частях Европы.
Эстонцы. Четыре года тому назад совместно с Центром переливания крови в Таллине было проведено исследование крови на предмет распределения ряда генов. У доноров эстонцев при этом использовались те же самые маркеры, что и у финнов. Очень жаль, что серия была небольшой, менее 500 человек, и касалась групп крови АВ0 и Rh. Выборка достаточно хорошая, хотя и не взвешена, как финская.
За исключением очень редких случаев, практически все маркеры, названные выше «типично финскими», представлены и в данной выборке, т. e. UIa, трансферрин B0-ll Dem, а также очень редкий Ъпп- Если сравнивать частоты маркеров в Финляндии и Эстонии, то можно сказать, что частоты изученных факторов в Эстонии представлены в меньшей концентрации, их частота составляет примерно половину значений этих частот, характерных для финнов (табл. 4).
Таблица Присутствие некоторых генетических маркеров в Финляндии и Эстонии Гены Генные частоты Гены Генные частоты Финляндия Эстония Финляндия Эстония а2 0,10 Около 0,07 UI" 0,012 0,005 м 0,64 0,59 Tf В0 -1 0,012 0,008 Fya 0,47 0,40 Tf Dvhi 0,009 0,005 К 0,02 0,25 Di" 0,0 0,003Другие маркеры, встречающиеся с большей частотой в Финляндии, по-видимому, распространяются сходным образом. Частота М, меньшая 0,60 (0,5918), и частоты Duffy и Gc соответствуют частотам, свойственным другим европейским популяциям. С точки зрения уже приводившихся доводов, сравнить частоты АВО и Rh нельзя. Все же отношение ?2'.A среди финнов исключительно высокое, отчетливо ниже или почти равно 1:3, в противоположность этому у эстонцев оно близко к 1 : 2.
Среди эстонцев были найдены два носителя группы крови Diego. Поскольку до сих пор эта группа крови не была выявлена у финнов, ее наличие может свидетельствовать о том, что фактор Diego мог быть приобретен от ранних финских поселенцев. Необходимо отметить, что D" также является чрезвычайно «экзотическим» маркером, он до сих пор не был обнаружен у европейцев.
Небольшой размер выборки требует осторожности при оценке результатов, представленных выше. Однако очевидно, что эстонский генофонд подобен генофонду финнов, хотя почти на 50% «разведен» генами, заимствованными от окружающих популяций, вероятно представляющих славянские и германские элементы, что находится в соответствии с известными историческими фактами. Территория Эстонии в прошлом служила ареной многих войн, испытала гнет феодализма, чего пе было па севере, в Финляндии. Это означает, что при исследовании более ранних фаз формирования балтийских финнов в качестве эталонной популяции должны скорее использоваться финны, чем их потомки по Прямой линии — эстонцы. Вот почему было затрачено столько времени на оценку сохранности естественного генетического пула нашей изолированной пограничной популяции.
Венгры. О близости финнов и эстонцев свидетельствуют лингвистические и археологические данные. К тому же со времена расхождения этих групп прошло менее 2 тыс. лет или примерно 50 поколений. Исторический разрыв между балтийскими финнами и венграми гораздо больше, за это время могли произойти многие события, имеющие значение для генетики исследуемых популяций.
В этой связи необходимо упомянуть об одной типичной черте угро-финской лингвистической группы. В сравнении с другими лингвистическими семьями финно-угорские языки распространены на небольшой территории. Таким образом, мы говорим, по-видимому, о братских нациях, расстояние между которыми, но крайней мере языковое, намного меньше, чем между большинством других европейских национальностей.
Совместно с Институтом гематологии в Будапеште мы недавно провели исследование частоты генов у 500 венгров из различных районов Венгрии. Были прослежены места рождения родителей и была сделана попытка отобрать только «чистых» представителей венгров. В данной работе не буду детально анализировать результаты, но сделаю несколько замечаний, представляющих интерес.
Первое касается гомогенности изученной популяции. По-видимому, венгры не образуют гомогенной популяции с равномерным распространением генов, как у финнов. Локальные различия значительны и, очевидно, необходимо изучить ряд отдельных хорошо взвешенных выборок, чтобы получить достоверные результаты в отношении всей популяции. Этот факт иллюстрирует распределение редкого маркера — трапсферрина В0. Было выявлено, что все индивидуумы с этим маркером проживали в западной части страны.
Что касается собственно финских маркеров, то за исключением одного они отсутствуют в исследованной выборке. Только у одного человека был найден ген Tf В0-, но и этот случай демонстрирует генетическую связь между венграми и финнами.
Таблица 5. Частота некоторых маркирующих генов в Финляндии Венгрии Гены Венгрии Финляндия Гены Венгрия Финляндия 0,057 0,096 та 0,00 0,01 м 0,64 0,56 Gcx 0,80 0,68 rh (cde) 0,34 0,41 Tf вл_1 0,00 0,001 Kell 0,036 0,002 Tf Dchi 0,009 0,000 Fya 0,39 0,47 Tf I)Fin 0,0005 0,000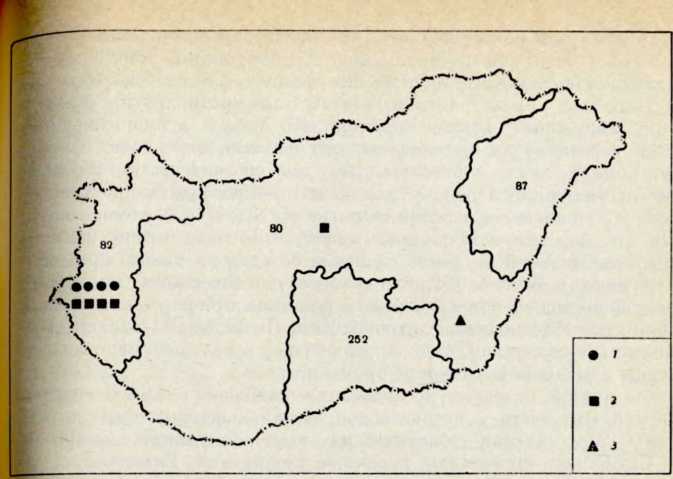
Рис. 3. Типы трансферрииов В0С, ВгС и Вд^С на территории Венгрии 1 — В„С, г — ВгС, 3 — Bo_iC. Цифрами обозначено число обследованных лиц
Хотя связь все же существенная, необходимо помнить, что до сих пор этот маркер был обнаружен только у индейцев навахо, но не среди исследованных азиатских популяций. Таким образом, значение этой скудной находки могло бы быть большим, если бы вместо В0-1 был обнаружен другой типичный маркер, Dcm, который распространен среди монгольских популяций (рис. 3).
Частота генетических маркеров в Венгрии не обнаруживает ничего такого, что указывало бы на общий с финнами генофонд (табл. 5). Частота Аг низкая, частота Bk такая же, как и в других частях Европы, это же относится и к Kell, Duffy, различным группам сыворотки и т. д.
Однако необходимо помнить, что размер выборки ограничен, а период разрыва времени с момента разделения финнов и венгров много более 2 тыс. лет. С другой стороны, то, что мы не смогли продемонстрировать явной генетической связи между этими двумя популяциями, не слишком удивительно. Как было показано, «разведение» генофонда балтийских финнов на протяжении всего лишь нескольких столетий составило около 50%. Соответствующее «разведение», которое определенно имело место Как у венгров, так и у балтийских финнов, должно быть у первых во много раз больше, скажем, даже больше 90%. «Разведение» подобного типа делает любые попытки продемонстрировать Генетические связи почти невозможными и поэтому бесполезными.
Суммируя, можно сказать, что представленные данные показывают, что генетическая модель популяций Финляндии, а возможно, также и балтийских финнов — имеет своеобразные и типичные характеристики, неизвестные среди других европейских популяции. Также очевидно, что финны лучше сохранили свой первоначальный генофонд, чем потомки балтийских финнов по прямой линии — эстонцы. Эти два обстоятельства образуют почти уникальную основу для попыток проследить происхождение угро-финской языковой семьи и исследовать ее гомогенность, До сих пор результаты показывали, что связь между ранними и поздними ветвями очень слабая и ее едва ли можно продемонстрировать. Иначе обстоит дело с генетическими маркерами, возможно менее подверженными влиянию отбора, чем признаки, принятые в физической антропологии. Последние вызывают определенный пессимизм, так как, вероятно, в большей мере подвержены изменениям в течение времени.
С другой стороны, исследования наиболее поздних передвижений населения и потока генов дают обнадеживающие результаты. Составление генетических карт популяций, например, в СССР дает интересные и ценные результаты. Естественно, что нас особенно интересуют характеристики популяций, живших или живущих по соседству с нами.
Более 800 000 книг и аудиокниг! 📚
Получи 2 месяца Литрес Подписки в подарок и наслаждайся неограниченным чтением
ПОЛУЧИТЬ ПОДАРОК