Глава 7 Посланцы генов
В главе 6 была описана двуспиральная модель ДНК. Мы узнали, как с ее помощью удалось объяснить правила спаривания оснований и рентгенографические данные. Вслед за тем мы увидели, что эта модель автоматически подсказывает нам замечательно простую схему репликации ДНК — процесса, которым сопровождается появление каждого нового поколения. Естественно предположить, что наследственная информация, передаваемая от поколения к поколению, содержится в последовательности основания ДНК, поскольку в остальном молекула ДНК по всей своей длине одинакова, элементы главной цепи ДНК повторяются без каких бы то ни было вариаций. Мы вынуждены предположить, что последовательность оснований А, Г, Ц и Т носит характер закодированного сообщения, которое и заключает в себе наследственную информацию. Проблему конкретных особенностей кода мы отложим до следующей главы. Пока что будем просто считать, что какой-то код действительно существует, а обсуждать будем другую проблему — проблему выражения закодированной информации. Нас будет интересовать способ, посредством которого наследственная информация управляет развитием клеток потомства, — способ воплощения в потомстве совокупности признаков, определяющих данный вид.
Принимая во внимание гипотезу «один ген — один фермент», а также учитывая то обстоятельство, что ДНК содержит линейную последовательность оснований, а белок представляет собой линейную последовательность аминокислот, мы можем сформулировать проблему иначе: как последовательность оснований ДНК, содержащейся в хромосомах, преобразуется в последовательность аминокислот множества различных белков, которых в любой клетке насчитывается несколько тысяч?
Прежде чем углубиться в обсуждаемую здесь проблему, я должен буду познакомить вас еще с одним видом нуклеиновых кислот — с рибонуклеиновой кислотой (РНК). Я сожалею, что вынужден осложнять вам жизнь, но что делать? Как я уже говорил в самом начале, жизнь действительно сложна, и мы должны принимать ее такой, как она есть.
РНК (рис. XII) очень похожа на ДНК. Она отличается от ДНК только в двух отношениях. Во-первых, сахар у нее несколько иной — вместо дезоксирибозы она содержит рибозу, но это различие очень невелико, речь идет всего-навсего об одной гидроксильной группе (ОН) каждого сахарного кольца.
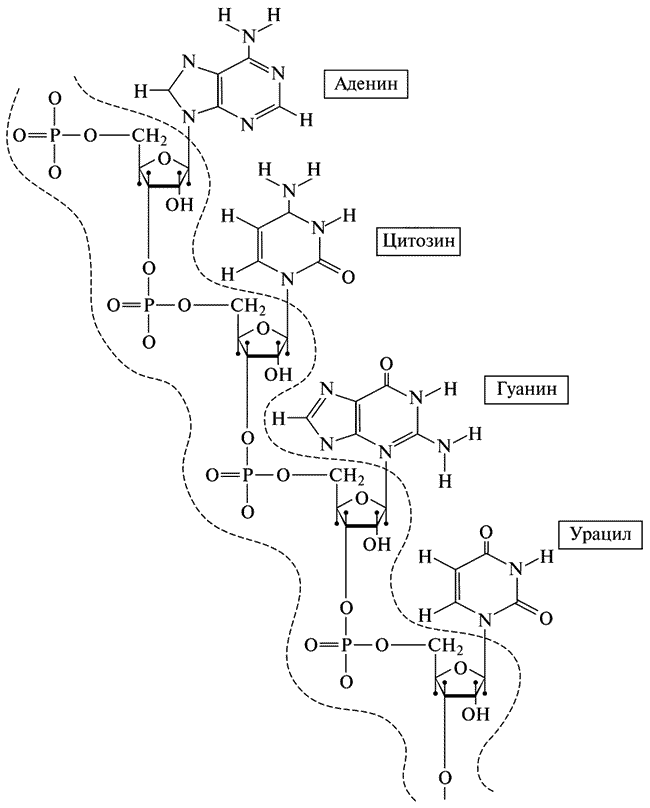
Рис. XII. Структура рибонуклеиновой кислоты (РНК).
Watson J.D. Molecular Biology of the Gene, New York, Benjamin, Inc., 1965, p. 91 (Дж. Уотсон, Молекулярная биология гена, «Мир», М., 1967).
Вторая особенность состоит в том, что набор оснований в РНК несколько иной, чем в ДНК. Вместо тимина в РНК появляется новое основание — урацил (сокращенно У). Итак, в РНК содержатся основания А, Г, Ц и У. Урацил химически очень близок тимину; он также относится к классу пиримидинов (с одним кольцом) и, подобно тимину, составляет пару с аденином.
Теперь снова вернемся к синтезу белка. Сравнительно недавно было показано, что в клетке синтез белков в основном происходит не в ядре, где содержатся хромосомы и, следовательно, ДНК, а в цитоплазме. А если так, то ДНК не может непосредственно управлять синтезом белков, поскольку она не там расположена: в ходе синтеза белков ДНК ядро не покидает. Мы вынуждены заключить, что существует какой-то механизм внутриклеточной передачи информации: инструкция, закодированная в ДНК, должна быть передана из ядра в цитоплазму — туда, где идет синтез белка. Недавно выяснилось, что посланцы генов, т. е. молекулы, ответственные за внутриклеточную транспортировку информации и за преобразование этой информации в последовательность аминокислот, — это опять-таки молекулы нуклеиновой кислоты, но уже иной. Я имею в виду РНК, с которой мы только-только познакомились. Как мы помним, передача информации между клетками осуществляется с помощью ДНК.
Существует несколько разновидностей РНК. Мы в этой главе рассмотрим три из них: рибосомную РНК, транспортную РНК и информационную РНК, которую также называют матричной РНК и РНК-посредником.
Теперь посмотрим, где же в действительности синтезируется белок. Местом синтеза служат крошечные частицы — рибосомы, которые в огромном количестве присутствуют в большинстве живых клеток. Обычно они прикреплены к мембранам — тонким перегородкам, образующим внутри цитоплазмы (т. е. вне ядра) густую сеть. Уже довольно давно известно, что белок синтезируется именно в рибосомах. Если клетку мы уподобляем фабрике по производству белков, то рибосомы на этой фабрике выполняют роль сборочных конвейеров. Можно выделить рибосомы из клетки, поставить дополнительное оборудование (ферменты), обеспечить их сырьем и энергией, и тогда их можно будет заставить вести синтез белка в пробирке, однако при том обязательном условии, что к ним поступает нужная информация. Описанная здесь так называемая бесклеточная система оказалась ценнейшим орудием исследования, позволявшим пролить свет на механизм биосинтеза белков.
Рибосомы построены из белка и РНК; эту РНК называют рибосомной. Нужно сразу честно признаться, что мы до сих пор не знаем, зачем она нужна. Одно время думали, что рибосомная РНК как раз и служит матрицей для синтеза белка. Это означало бы, что код рибосомной РНК соответствует последовательности аминокислот в белке, который синтезируется на данной рибосоме. Однако оказалось, что не так это все просто. Возможно, рибосомная РНК выполняет какую-то структурную роль. Во всяком случае, мы теперь твердо знаем, что любая рибосома при наличии нужной информации может делать любой белок, причем информация всегда поступает извне и с самой рибосомой не связана.
Информацию переносит РНК другой из упомянутых разновидностей, а именно информационная РНК (или РНК-посредник). Функция этого переносчика информации состоит в том, чтобы извлекать информацию оттуда, где она хранится, и доставлять ее туда, где она используется. Информация извлекается из расположенной внутри ядра ДНК и доставляется в цитоплазму — к рибосомам. Информационная РНК была открыта совсем недавно в результате целого ряда очень остроумных экспериментов, о которых, будь у меня больше места, я бы с удовольствием рассказал. Во многих клетках информационная РНК, по-видимому, весьма неустойчива. Очевидно, в большинстве случаев, как только она выполнит свою работу, сделав несколько молекул белка, она тут же рассыпается на части. Во всяком случае, в каждый данный момент времени количество информационной РНК внутри клетки весьма незначительно. Вот почему даже тогда, когда уже знали, что подобный переносчик информации должен существовать, обнаружить его было очень трудно.
Вся цепь событий, о которых я сейчас собираюсь рассказать, была установлена в результате очень сложных экспериментов, и я простоты ради сразу опишу весь механизм, не останавливаясь на том, каким способом эти сведения были добыты.
Механизм этот поистине замечателен. Информационная РНК образуется в ядре. И строится она так, что ее основания подбираются в соответствии с последовательностью оснований одной из цепей хромосомной ДНК, иными словами, подбираются согласно правилам Чаргаффа. Детали процесса нам не известны, но мы знаем, что он существует на самом деле. Итак, в согласии с правилами Чаргаффа, вставляя всякий раз в пару с аденином урацил вместо тимина, мы получим цепь информационной РНК, последовательность оснований в которой комплементарна одной из цепей ДНК[240]. Одна молекула информационной РНК может соответствовать одной или нескольким (очень немногим) молекулам белка. Как только цепь РНК построена, она переходит в цитоплазму и блуждает там до тех пор, пока не найдет какую-нибудь рибосому, к которой она и прикрепляется.
Дальше необходимо, чтобы к рибосомам были доставлены аминокислоты, из которых должна строиться полипептидная цепь. Они, конечно, должны быть выстроены в определенном порядке в соответствии с инструкцией, закодированной в информационной РНК. Теперь мы оказываемся перед лицом одной трудности логического порядка: цепочка оснований нуклеиновой кислоты способна «узнавать» другую цепочку оснований (как это происходит при репликации ДНК), но будет игнорировать цепочку аминокислот. Легко понять, как основания «узнают» друг друга — на то и существуют правила спаривания. Но как представить себе химический механизм, с помощью которого последовательность оснований могла бы «узнать» аминокислоту? Для того чтобы найти выход из этого тупика, была выдвинута — задолго до своего подтверждения — гипотеза о существовании специальной адапторной молекулы, которая, так сказать, с одного конца распознает последовательность из нескольких оснований цепи РНК, а другим концом может специфически связывать нужную аминокислоту. Предсказание подтвердилось: адаптор нашли, и оказалось, что это также РНК, а именно транспортная РНК (ее еще называют адапторной или растворимой РНК).
Транспортная РНК отличается от остальных РНК, с которыми мы встречались, тем, что ее молекулы значительно меньше. Они содержат всего по 70–80 оснований. Где-то среди них расположена последовательность оснований, комплементарная определенной последовательности оснований в информационной РНК (иначе говоря, эта последовательность транспортной РНК способна «узнавать» соответствующий участок в последовательности информационной РНК). А где-то в другом конце молекулы находится участок, способный «узнавать» определенную аминокислоту.
Отсюда следует, что должно существовать как минимум двадцать разных сортов транспортной РНК — по одной на каждую из двадцати аминокислот. Еще это означает, что имеется двадцать специфических ферментов, управляющих присоединением каждой из аминокислот к своей транспортной РНК. Дело в том, что реакции между транспортной РНК и аминокислотой, как и большинство других реакций, протекающих в живой клетке, в отсутствие фермента не идут. Сразу скажем, что все двадцать видов транспортных РНК и все ферменты были действительно обнаружены в клетке.
В ядре на ДНК формируется информационная РНК. Она поступает в цитоплазму и прикрепляется к рибосоме, а затем за нее цепляются молекулы транспортных РНК, несущие каждая свою аминокислоту. В результате аминокислоты выстраиваются в том порядке, который диктуется последовательностью оснований в РНК-посреднике. Теперь представьте себе, что аминокислоты соединяются друг с другом, и полипептидная цепь готова.
Эта схема может показаться сложной, но согласитесь, что она очень остроумна. В действительности она еще сложнее, чем я вам рассказал; многие детали нам до сих пор неизвестны. Это и неудивительно, ведь вся эта область исследований очень молода, и даже сама информационная РНК была впервые обнаружена только в 1960 году. Я упомянул здесь еще лишь о двух усложнениях.
Прежде всего, структура самих рибосом далеко не проста. Они построены из двух субъединиц разного размера — одна побольше, другая поменьше. А почему это так, мы имеем пока самые смутные представления.
Далее было обнаружено, что рибосомы, активно участвующие в синтезе белка, всегда оказываются связанными в группы по пять-шесть штук, причем все они прикреплены к одной цепи информационной РНК. Эти комплексные структуры, названные полисомами, как раз и служат подлинными производителями белка. Можно отделить полисомы от свободных, не объединенных в группы рибосом, и тогда обнаруживается, что синтез белка идет именно в полисомах.
На электронной фотографии полисом явственно видно, как рибосомы связаны очень тонкой нитью. Причем есть все основания считать ее нитью информационной РНК. Здесь мы прямо на фотографии, так сказать во плоти, видим ту удивительную молекулу, существование которой первоначально было выведено всего лишь как неизбежное логическое следствие того факта, что синтезы белка и хромосом в клетке разобщены. Создается впечатление, что одна молекула информационной РНК участвует одновременно в синтезе нескольких молекул белка. Рибосома прикрепляется к информационной РНК с одного конца и «прокладывается» вдоль по цепи до другого конца. Достигнув дальнего конца, она соскальзывает с цепи, и в этот момент в раствор освобождается вновь синтезированная молекула белка.
Суть всей схемы заключается в том, что биологическая информация одномерна и может быть записана в виде линейной последовательности. Полипептидная цепь белка представляет собой линейную последовательность аминокислот. Информация записана в нуклеиновых кислотах — как в ДНК, так и в информационной РНК, подобно строке в книге. Однако, как мы помним, большинство белков являются трехмерными объектами, их полипептидные цепи свернуты очень сложным и специфическим образом.
Как же происходит свертывание цепи миоглобина? До сих пор в нашей схеме белку было предоставлено только одно измерение, мы рассматривали его лишь как последовательность аминокислот. Теперь надо решать, является ли свертывание цепи самопроизвольным процессом или клетка содержит информацию не только о последовательности аминокислот в полипептидной цепи, но также о способе ее свертывания по окончании синтеза. Можно было бы вообразить, что в клетке имеются какие-то специальные трехмерные матрицы — своего рода формы для изготовления трехмерных молекул белка. Но пока что в пользу их существования нет абсолютно никаких данных, хотя таких матриц в каждой клетке должно было бы быть столько же, сколько и белков, т. е. порядка нескольких тысяч. Да и в самом деле едва ли можно себе представить, как такая система могла бы работать. Посмотрите только на модель белка и вам станет ясно, какие сложнейшие проблемы здесь возникают. Взять хотя бы то, что такую модель просто-напросто нельзя было бы извлечь из формы.
Сейчас принято считать, хотя это и не доказано, что белки, синтезированные в виде линейной последовательности, свертываются сами. Иными словами, сложная пространственная конфигурация молекулы белка возникает самопроизвольно. Теперь эта простая гипотеза выглядит еще более правдоподобно, поскольку недавно было показано, что молекулы белка (фермента) с искусственно развернутыми цепями, даже в пробирке, где нет ни рибосом, ни нуклеиновых кислот (вообще никаких других компонентов живой клетки), могут снова свертываться нужным образом, так что уже через несколько минут активность фермента восстанавливается.
Тот факт, что полипептидные цепи свертываются самопроизвольно, придает изучению трехмерной структуры белков еще больший интерес, поскольку у нас, таким образом, появляется надежда выяснить правила свертывания цепей с известной последовательностью аминокислот. В дальнейшем, пользуясь этими правилами, мы смогли бы установить трехмерную структуру белка по его аминокислотной последовательности, минуя все тяготы, связанные с применением рентгеноструктурного анализа. Впрочем, я думаю, что пока нам до этого еще очень и очень далеко.
Подведем итоги. Мы описали схему, согласно которой информация, хранимая в ядерной ДНК, считывается в цепь информационной РНК. Эта РНК поступает в цитоплазму, и там в ней прикрепляется одна или несколько рибосом. Рибосомы перемещаются вдоль цепей РНК и по мере своего движения синтезируют белок. Аминокислоты доставляются к этому конвейеру молекулами транспортной РНК и размещаются здесь в определенной последовательности в соответствии с кодом. Расшифровкой этого кода опять-таки ведают молекулы транспортных РНК: место, предназначенное для их аминокислоты, они распознают, отыскивая соответствующий участок в цепи информационной РНК.
Когда в главе 5 мы впервые заговорили о нуклеиновых кислотах как о носителях наследственных признаков любой живой клетки, я сказал, что подобную роль они могут играть только при условии, что им будут присущи три главных свойства: способность к репликации, способность к хранению информации и способность к управлению синтезом белков. Мы уже обсудили первое и третье свойства. В следующей главе пойдет речь о втором свойстве. Нас будет интересовать соответствие между последовательностью оснований в цепи нуклеиновой кислоты и последовательностью аминокислот в синтезируемой под контролем нуклеиновой кислоты цепи белка. Это и есть так называемая проблема кода. В самое последнее время в этой области были достигнуты огромные успехи, так что у нас в руках имеется почти полное решение проблемы, а ведь всего несколько лет назад она казалась невероятно трудной — никто даже и не надеялся, что узнать это решение суждено уже нашему поколению.
Более 800 000 книг и аудиокниг! 📚
Получи 2 месяца Литрес Подписки в подарок и наслаждайся неограниченным чтением
ПОЛУЧИТЬ ПОДАРОК