Глава 6 Нуклеиновые кислоты — молекулы наследственности
В главе 5 мы установили, что наследственная информация, хранящаяся в каждой клетке и передаваемая от поколения к поколению, заключена в клеточном ядре, в хромосомах. Элементы информации, ответственные каждый за синтез одного определенного белка, называются генами. Хромосомы состоят из белка и нуклеиновой кислоты (ДНК), но непосредственным носителем информации служат только ДНК. Мы познакомились с химической формулой ДНК: молекула ДНК представляет собой длинную цепь чередующихся сахарных и фосфатных групп, причем к каждой сахарной группе прикреплена еще боковая цепь — одно из четырех возможных оснований. Точно так же, как химическая формула белка мало что говорит о его функции, так и формула нуклеиновой кислоты представляет собой недостаточно полный символ, по которому нельзя судить, как проходит самовоспроизведение ДНК и как хранится информация.
И снова, как и раньше, естественным образом возникает вопрос: каково же взаимное пространственное расположение атомов в молекуле ДНК? Здесь опять-таки приходится обращаться за помощью к рентгеновской кристаллографии. В волокнах очищенной ДНК длинные молекулы ориентированы, как стебельки в пучке соломы. Примерно так же упакованы длинные тонкие молекулы кератина волос. Значит, если волокно ДНК поместить на пути пучка рентгеновских лучей, то должна получиться рентгенограмма, сходная по характеру с рентгенограммой волоса. Первая рентгенограмма ДНК была получена Астбюри в середине тридцатых годов [ХХ в.]. Снимок получился сильно размытым. Конечно, определить по такому снимку структуру ДНК — задача совершенно безнадежная. Тем не менее, и сам Астбюри, и другие исследователи пытались это сделать. И хотя некоторые из них (особенно Астбюри), как потом выяснилось, находились на верном пути, сделанные ими выводы были неубедительны, и проблема долгие годы оставалась нерешенной.
Так продолжалось до начала пятидесятых годов. Примерно к этому времени относятся два очень важных экспериментальных достижения, заставивших совсем по-иному смотреть на структуру и роль ДНК.
Первое из них связано с именем Э. Чаргаффа. Вы помните, что тогда почти ничего не знали о количественном соотношении между основаниями, входящими в состав ДНК; считалось даже, что они представлены в равном количестве. Заслуга Чаргаффа состоит в том, что он первый получил по-настоящему чистые образцы ДНК и сделал очень аккуратный анализ относительного числа оснований в каждом образце. Он обнаружил, что процентное содержание четырех оснований — А, Г, Ц и Т — от вида к виду довольно сильно меняется, так что, вообще говоря, содержание в ДНК разных оснований далеко не одинаково. Кроме того, были открыты две весьма замечательные закономерности, которые вместе составляют так называемые правила спаривания оснований (правила Чаргаффа): из какого бы организма ни была выделена ДНК и как бы сильно процентный состав оснований ни отличался от предполагавшегося ранее отношения 1:1:1:1:1, количество А всегда равно количеству Т, а количество Г — количеству Ц. Вы, должно быть, помните, что А и Г — это пурины, содержащие два кольца атомов, а Ц и Т — пиримидины, содержащие по одному кольцу. Следовательно, равными оказываются количества «больших» и «малых» оснований: число А («большие» основания) равно числу Т («малые» основания); число Г («большие») равно числу Ц («малые»). В то время открытые закономерности были необъяснимы, но в конечном счете именно они послужили ключом к разгадке структуры ДНК.
Вторым не менее важным достижением мы обязаны М. Уилкинсу и Р. Франклин совместно с их сотрудниками. Им удалось сильно повысить качество рентгенограмм, получаемых от волокон ДНК. В этом вы сами можете легко убедиться. Сходство двух рентгенограмм не вызывает сомнений, но на новом снимке на месте размытых пятен имеется много четких рефлексов. С таким снимком кристаллографы вполне согласятся работать.
Таковы были эти новые экспериментальные факты. Ими блестяще воспользовались Дж. Уотсон и Ф. Крик. Уотсон, которому в ту пору (1953 г.) было только 24 года, приехал из Америки в Кембридж, в нашу лабораторию, и должен был вместе со мной заниматься изучением структуры белка. Но белком он так почти и не занимался. Как он сам первый признал, это была для него чересчур тяжелая работа. Я, конечно, на этом потерял, но зато много выиграла биология в целом. Крик приехал в нашу лабораторию чуть раньше, и они с Уотсоном подолгу беседовали о том, как важно было бы расшифровать структуру ДНК. Они разглядывали последние рентгенограммы ДНК, обсуждали возможный смысл открытых Чаргаффом правил спаривания оснований, перепробовали чуть ли не все возможные виды моделей. В результате по прошествии всего нескольких недель после одной-двух неудачных попыток разгадка по существу была уже готова! Я даже затрудняюсь объяснить, как они ее нашли, — по-моему, и сами они едва ли смогли бы объяснить. Это было одно из тех прозрений, которые время от времени случаются в науке. Вы можете называть это гениальностью, вдохновеньем, как хотите. Одно можно сказать: до 1953 года такое прозрение было бы невозможно, поскольку оно целиком зависело от открытия правил спаривания оснований, а также от информации, содержащейся в улучшенных рентгенограммах. Но, как только все эти сведения были добыты, ответ удалось найти в поразительно короткий срок.
Модель структуры ДНК, построенная Уостсоном и Криком, — это двойная спираль; как показано на модели, молекулы ДНК состоят из двух цепей, идущих в противоположных направлениях и закрученных одна вокруг другой наподобие электрических проводов. В результате получается структура, немного напоминающая винтовую лестницу (рис. IX).
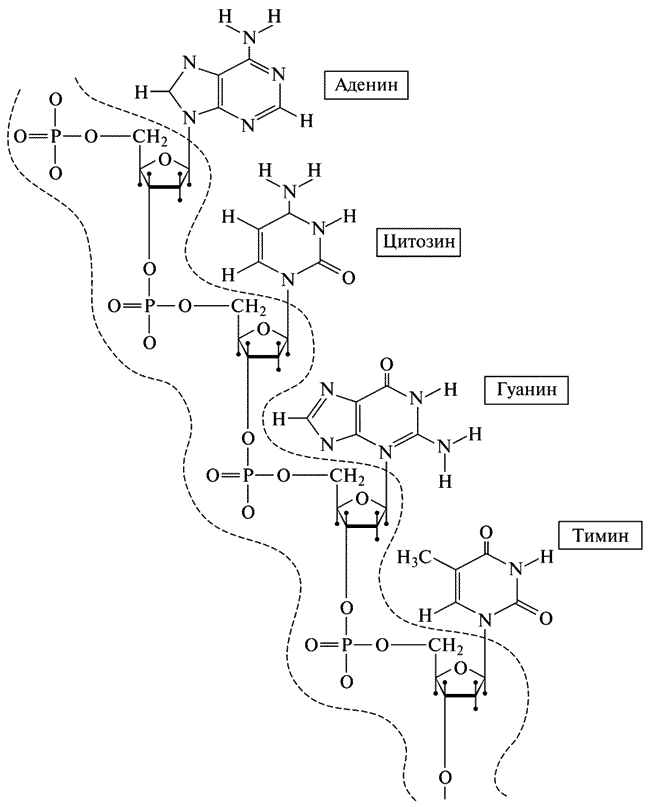
Рис. IX. Структура одноцепочной дезоксирибонуклеиновой кислоты (ДНК):
Главная цепь (остов) ограничена двумя волнистыми линиями. Вправо отходят боковые цепи. Всего возможны четыре разновидности боковых цепей, которые чередуются в совершенно нерегулярной последовательности. Watson J. D. Molecular Biology of the Gene, New York, Benjamin, Inc., 1965, p. 91 (Дж. Уотсон, Молекулярная биология гена, «Мир», М., 1967).
Ступеньками ее служат пары оснований, скрепленных теми же слабыми водородными связями, с которыми мы уже встречались в альфа-спирали — там, где речь шла о структуре белка. Наиболее существенно в структуре ДНК то, что каждая пара оснований, составляющая ступеньку, обязательно содержит одно большое и одно малое основания, так что встречаются только пары А-Т и Г-Ц. Ступенька не может состоять из двух малых оснований, скажем из Т и Ц, двух Т или двух Ц, так как они не достанут до середины и не смогут соединиться водородной связью. Не может она состоять и из двух больших оснований — А и Г, А и А или Г и Г — не хватит места. Нельзя соединить в пары также А с Ц или Г с Т — это невозможно просто потому, что их химическая структура исключает возможность образования подходящих водородных связей. Таким образом, правила Чаргаффа получают простое объяснение, исходя из структуры: на каждой ступеньке должна быть либо пара А-Т, либо пара Г-Ц, и, следовательно, общее число А должно равняться общему числу Т, а число всех Г — числу всех Ц. Объяснение получилось настолько естественным, что каждый, кто знакомится с деталями предложенной структуры, а в особенности кристаллографы и химики-структурщики, у которых уже наметан глаз на такого типа модели, сразу получают полное удовлетворение. Возникает такое ощущение, будто иначе и не может быть.
Имея в руках модель, подобную двойной спирали Уотсона-Крика, вы можете проверять ее тысячами способов. Вы можете рассчитать, какую она должна давать рентгенограмму, и сравнить, насколько результат соответствует экспериментально полученной рентгенограмме. Используя метод проб и ошибок, Уотсон и Крик впоследствии слегка изменили свою модель и получили достаточно удовлетворительное соответствие с данными рентгеноструктурного анализа, а позднее Уилкинс внес еще кое-какие уточнения, так что теперь соответствие просто превосходное. Но ни одна из внесенных поправок не нарушила великой простоты первоначальной модели. Структура Уитсона-Крика подтверждена теперь во всех подробностях, и мы можем быть совершенно уверены, что в волокнах ДНК расположение цепей носит точно такой характер.
Однако ДНК в волокнах совсем не обязательно должна быть тем же, чем она является в живых клетках. Заранее нельзя сказать, что структура, однозначно установленная для ДНК внутри волокон, описывает состояние молекул ДНК в их биологическом окружении. В последние несколько лет многие биохимики и физико-химики изучали ДНК уже непосредственно в живых клетках, такой, как она есть. За недостатком места мы не будем описывать все эти методы. Скажу только результат: установлено, что обычно в живой клетке ДНК тоже присутствует в виде двойной спирали.
В наши дни электронный микроскоп позволяет непосредственно рассмотреть отдельные молекулы ДНК. Можно видеть короткий участок молекулы ДНК, имеющий в поперечнике две десятимиллионные доли сантиметра. Вот она, настоящая нить жизни. В том, что это действительно так, мы не замедлим убедиться. Сразу же возникает вопрос: каким образом она выполняет свою главную функцию — самовоспроизведение; каким образом получается, что после деления в каждой из дочерних клеток ДНК оказывается идентичной ДНК родительской клетки? Уотсон и Крик предложили смелую и вместе с тем очень простую гипотезу. Они допустили, что спираль ДНК расплетается на две одиночные цепи, а затем из нуклеотидов, свободно плавающих в клетке, формируется вдоль каждой цепи еще одна цепь, причем основания, соединяющиеся с основаниями старой цепи, подбираются в соответствии с правилами Чаргаффа. Легко видеть, что в конечном счете будут построены две одинаковые двойные спирали, идентичные первоначальной (рис. X).
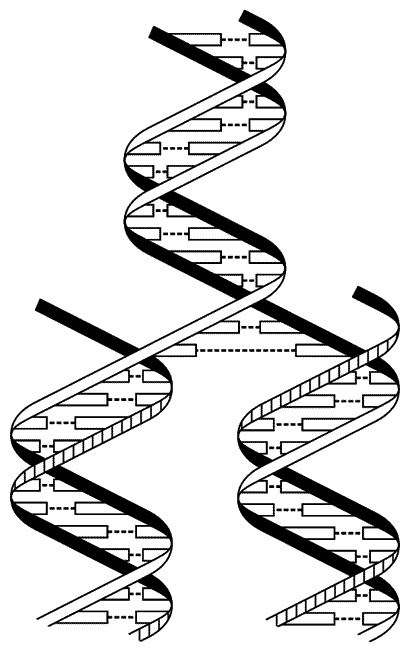
Схема репликации ДНК, согласно Уотсону-Крику:
Двойная спираль (в верхней части рисунка) раскручивается, и одновременно вокруг каждой из двух цепей исходной молекулы ДНК формируются новые цепи (внизу). В результате образуются две двойные спирали согласно правилам спаривания оснований, так что новые двойные спирали имеют ту же последовательность оснований, что и исходная молекула ДНК.
Этот процесс можно сравнить с печатанием фотокарточек с негатива. Различие здесь только в том, что в случае ДНК любую цепь можно рассматривать как «негатив» для другой, так что в этом смысле словно бы и нет разницы между позитивом и негативом. Совсем недавно с помощью электронного микроскопа удалось даже заснять ДНК в момент репликации.
Такова была гипотеза. Но биологи сразу заметили, что перед ней неизбежно встанет масса трудностей, проистекающих в первую очередь из громадной длины молекул ДНК. В одной клетке человека вся ДНК, распределенная в 46 хромосомах, содержит что-то около миллиарда пар оснований; ее полная длина достигает едва ли не метра. А если составить цепочку из ДНК всех клеток одного человека, то она, пожалуй, сможет протянуться через всю солнечную систему. И вот почти целый метр ДНК должен быть как-то свернут внутри одной клетки, размеры которой в поперечнике обычно составляют не более тысячной доли сантиметра. Если гипотеза Уотсона-Крика верна, то в процессе репликации вся эта ДНК должна быть раскручена на одиночные нити. Всякий, кто хотя бы раз пробовал расплести длинный шнур, состоящий из двух электрических проводов, сразу же поймет, почему биологи увидели здесь проблему: непонятно, что помешает еще до конца не расплетенной цепи спутаться в безнадежный клубок.
Поэтому было очень важно придумать такой решающий эксперимент, который мог бы служить проверкой предложенной гипотезы. Такой эксперимент задумали и успешно осуществили Меселсон и Сталь. Они выращивали определенный вид бактерий в искусственной среде, содержащей тяжелый азот (азот-15) — изотоп азота, атомы которого тяжелее атомов обычного азота (азот-14). Через некоторое время у этих бактерий весь азот в основаниях ДНК был представлен только тяжелым азотом. Как можно подсчитать, плотность такой ДНК должна почти на 1 % превышать плотность нормальной ДНК. Вырастив достаточное количество бактерий, содержащих тяжелую ДНК, Меселсон и Сталь переносили их на среду, содержащую обычный легкий азот (азот-14). На новой среде бактерии размножались путем деления, причем теперь для синтеза новой ДНК они могли использовать только легкий азот. Далее проводился анализ ДНК потомства. Что должно было получиться? Если гипотеза Уотсона-Крика верна, то в первом поколении потомство перенесенных бактерий должно содержать, так сказать, «полутяжелую» ДНК, которая легче, чем родительская, но тяжелее обычной, поскольку в каждом молекуле этой ДНК должна была присутствовать одна тяжелая и одна обычная цепь. Плотность такой ДНК должна примерно на 0,5 % превышать плотность нормальной ДНК. Выделенная ДНК будет представлять собой смесь ДНК двух плотностей. Половина всей ДНК должна иметь нормальную плотность, поскольку ее молекулы будут составлены из двух легких цепей, остальная ДНК будет «полутяжелой», т. е. содержащей по одной легкой и по одной тяжелой цепи.
Вот что предсказывает теория. А как проверить эти предсказания? Для этого надо найти чувствительный метод измерения плотности ДНК, который позволял бы обнаруживать различия в плотности ДНК, составляющие 0,5 или 1 %. Наиболее подходящим оказался метод градиентной колонки. Градиентная колонка — это колонка с жидкостью переменной плотности: на дне плотность жидкости наибольшая, на поверхности наименьшая. Если в такую колонку бросить какой-либо предмет, то он под действием силы тяжести будет погружаться в жидкость до тех пор, пока его средняя плотность не сравняется с плотностью непосредственно прилегающих к нему слоев жидкости. В принципе этим методом можно воспользоваться для того, чтобы отделить тяжелую ДНК от легкой: в колонке они должны располагаться на разной высоте. Но так как различия в плотности здесь очень малы, необходимо повысить чувствительность метода, насколько это возможно; на практике колонку переменой плотности создают путем быстрого вращения в центрифуге раствора какой-нибудь соли. Мы как бы создаем довольно высокую искусственную «силу тяжести». (Примерно так же тренируют космонавтов: чтобы приучить их к большим перегрузкам, их сажают в кресло, которое быстро вращается по кругу большого радиуса.) Соль при вращении стремится сместиться ближе к дну колонки, так что в этом направлении плотность раствора постепенно возрастает. Если в колонку добавить ДНК, то она сместится туда, где плотность раствора совпадает с ее собственной плотностью. Опыты Меселсона и Сталя: сначала показано, где в кювете центрифуги располагается тяжелая ДНК, выделенная из бактерий до их переноса на обычную среду. Бактерии первого поколения, выращенные на среде с легким азотом, содержат ДНК, состоящую наполовину из легких и наполовину из тяжелых цепей, а бактерии второго поколения содержат смесь такой «полутяжелой» и нормальной ДНК. Другими словами, поведение ДНК точно соответствует предсказаниям, сделанным на основе гипотезы Уотсона-Крика. Этот эксперимент по своему содержанию необычайно прост и дает совершенно однозначные результаты — классический пример решающего эксперимента! После того как эти результаты были опубликованы, подавляющее большинство биологов вынуждено было признать, что гипотеза Уотсона — Крика должна быть верна, и хотя до сих пор не вполне ясно, как происходит раскручивание ДНК, нет никаких сомнений, что предложенный механизм репликации отражает реальные события, происходящие в процессе клеточного деления.
На этом мы заканчиваем наш рассказ о первой функции ДНК — функции самовоспроизведения. Мы видели, как построенная Уотсоном и Криком двуспиральная модель ДНК позволила предсказать способ репликации, а потом мы убедились, что предсказание это подтверждается очень простым и вполне доказательным экспериментом. В следующих двух главах мы рассмотрим две другие функции ДНК. Во-первых, мы обсудим, как ДНК управляет синтезом белков, как наследственная информация преобразуется в структуру ферментов и других белков, синтезируемых в дочерних клетках. И во-вторых, мы постараемся понять, в каком виде информация хранится в самой ДНК, с помощью какого кода она там записана?
Более 800 000 книг и аудиокниг! 📚
Получи 2 месяца Литрес Подписки в подарок и наслаждайся неограниченным чтением
ПОЛУЧИТЬ ПОДАРОК